Funkcije provodnih tkiva cvjetnica. Mehanička i vodljiva biljna tkiva
Provodno tkivo sastoji se od živih ili mrtvih izduženih stanica koje izgledaju poput cjevčica.
Stabljike i listovi biljaka sadrže snopove vodljivog tkiva. Provodno tkivo sadrži žile i sitaste cjevčice.
Plovila- sekvencijalno povezane mrtve šuplje stanice, poprečne pregrade između njih nestaju. Preko sudova voda i u njoj otopljeni minerali iz korijena ulaze u stabljiku i lišće.
Sitaste cijevi - izdužene žive stanice bez jezgre povezane u seriju jedna s drugom. Kroz njih se organske tvari iz lišća (gdje su nastale) kreću u druge organe biljke.
Vodljiva tkanina osigurava transport vode s mineralima otopljenim u njoj.
Ovo tkivo tvori dva transportna sustava:
- prema gore(od korijena do lišća);
- prema dolje(od listova do svih ostalih dijelova biljaka).
Uzlazni transportni sustav sastoji se od traheida i žila (ksilem ili drvo), a žile su napredniji vodiči od traheida.
U silaznim sustavima prolazi tok vode s produktima fotosinteze sitaste cijevi(floem ili floem).
Ksilem i floem tvore vaskularno-vlaknaste snopove - "cirkulacijski sustav" biljke, koji ga potpuno prodire, povezujući ga u jednu cjelinu.
Znanstvenici vjeruju da je pojava tkiva povezana u povijesti Zemlje s pojavom biljaka na kopnu. Kada se dio biljke našao u zraku, a drugi dio (korijen) u tlu, postalo je potrebno vodu i mineralne soli dopremiti iz korijena u lišće, a organske tvari iz lišća u korijenje. Tako su se tijekom evolucije biljnog svijeta pojavile dvije vrste vodljivih tkiva - drvo i ličje.
Kroz drvo (kroz traheide i posude) voda s otopljenim mineralima se diže od korijena do lišća - to je vodoprovodna ili uzlazna struja. Kroz floem (kroz sitaste cijevi) organske tvari nastale u zelenom lišću teku do korijena i drugih organa biljke - to je silazna struja.
Edukativna tkanina
Obrazovno tkivo nalazi se u svim rastućim dijelovima biljke. Obrazovno tkivo sastoji se od stanica koje su sposobne dijeliti se tijekom cijelog života biljke. Stanice se ovdje vrlo brzo slažu jedna s drugom. Dijeljenjem stvaraju mnoge nove stanice, čime se osigurava rast biljke u duljinu i debljinu. Stanice koje se pojavljuju tijekom diobe obrazovnih tkiva zatim se transformiraju u stanice drugih biljnih tkiva.

To je primarno tkivo iz kojeg nastaju sva ostala biljna tkiva. Sastoji se od posebnih stanica sposobnih za višestruke diobe. Upravo te stanice čine embrij bilo koje biljke.
Ovo tkivo se zadržava u odrasloj biljci. Locirano je:
- na dnu korijenskog sustava i na vrhovima stabljika (osigurava rast biljke u visinu i razvoj korijenovog sustava) - apikalno obrazovno tkivo;
- unutar stabljike (osigurava rast biljke u širinu i zadebljanje) – bočno obrazovno tkivo.
Za razliku od drugih tkiva, citoplazma obrazovnog tkiva je deblja i gušća. Stanica ima dobro razvijene organele koji osiguravaju sintezu proteina. Jezgru karakterizira velika veličina. Masa jezgre i citoplazme održava se u stalnom omjeru. Povećanje jezgre signalizira početak procesa stanične diobe, koja se odvija mitozom za vegetativne dijelove biljaka i mejozom za sporogene meristeme.
Značaj i raznolikost provodnih tkiva
Vodljiva su tkiva najvažnija komponenta većine više biljke. Obvezna su strukturna komponenta vegetativnih i rasplodnih organa spornih i sjemenskih biljaka. Provodna tkiva zajedno sa staničnom stjenkom i međustaničnim prostorima, nekim stanicama glavnog parenhima i specijaliziranim prijenosnim stanicama čine provodni sustav koji osigurava daljinski i radijalni transport tvari. Zbog posebnog dizajna stanica i njihovog položaja u tijelu biljke, provodni sustav obavlja brojne, ali međusobno povezane funkcije:
1) kretanje vode i minerala koje korijenje apsorbira iz tla, kao i organskih tvari nastalih u korijenju, u stabljiku, lišće i reproduktivne organe;
2) kretanje produkata fotosinteze od zelenih dijelova biljke do mjesta njihove upotrebe i skladištenja: korijenje, stabljike, plodovi i sjemenke;
3) kretanje fitohormona kroz biljku, čime se stvara određena njihova ravnoteža, koja određuje brzinu rasta i razvoja vegetativnih i reproduktivnih organa biljaka;
4) radijalni transport tvari iz provodnih tkiva u obližnje žive stanice drugih tkiva, na primjer, u asimilirajuće stanice mezofila lista i stanice meristema koje se dijele. U njemu mogu sudjelovati i parenhimske stanice medularnih zraka drva i kore. Transmisivne stanice s brojnim izbočinama stanične membrane, smještene između provodnog i parenhimskog tkiva, imaju veliku važnost u radijalnom transportu;
5) vodljiva tkiva povećavaju otpornost biljnih organa na deformirajuća opterećenja;
6) provodna tkiva čine kontinuirani razgranati sustav koji povezuje biljne organe u jedinstvenu cjelinu;
Pojava provodnih tkiva rezultat je evolucijskih strukturnih transformacija povezanih s izlaskom biljaka na kopno i odvajanjem njihova zraka i ishrana tla. Najstarija provodna tkiva, traheide, pronađena su u fosilnim rinofitima. Najviši razvoj dosegli su u modernim angiospermama.
Tijekom procesa individualnog razvoja iz prokambija nastaju primarna provodna tkiva na mjestima rasta sjemenog zametka i pupoljaka obnove. Sekundarna provodna tkiva, karakteristična za dikotiledone angiosperme, stvara kambij.
Ovisno o funkcijama koje obavljaju, provodna tkiva se dijele na tkiva uzlazne struje i tkiva silazne struje. Glavna svrha uzlaznog tkiva je prijenos vode i u njoj otopljenih minerala od korijena do viših nadzemnih organa. Osim toga, kroz njih se kreću organske tvari nastale u korijenu i stabljici, kao što su organske kiseline, ugljikohidrati i fitohormoni. Međutim, izraz "uzlazna struja" ne treba jednoznačno shvatiti kao kretanje odozdo prema gore. Uzlazna tkiva osiguravaju protok tvari u smjeru od usisne zone do vrha izdanka. U tom slučaju transportirane tvari koristi kako sam korijen, tako i stabljika, grane, lišće, reproduktivni organi, bez obzira nalaze li se iznad ili ispod razine korijena. Na primjer, kod krumpira voda i elementi mineralne prehrane teku kroz uzlazna tkiva u stolone i gomolje formirane u tlu, kao iu nadzemne organe.
Tkanine silazna struja osigurati odljev produkata fotosinteze u rastuće dijelove biljaka i organe za pohranu. U ovom slučaju prostorni položaj fotosintetskih organa nije bitan. Na primjer, u pšenici, organske tvari ulaze u razvoj zrna iz listova različitih slojeva. Stoga nazive "uzlaznih" i "silaznih" tkanina treba tretirati kao ništa više od utvrđene tradicije.
Provodna tkiva uzlazne struje
U uzlazna tkiva spadaju traheide i žile (traheje), koje se nalaze u drvenastom (ksilemskom) dijelu biljnih organa. U tim tkivima kretanje vode i u njoj otopljenih tvari odvija se pasivno pod utjecajem pritiska korijena i isparavanja vode s površine biljke.
Traheidi su starijeg porijekla. Nalaze se u višim spornim biljkama, golosjemenjačama, a rjeđe u kritosjemenjačama. Kod angiospermi su tipične za najmanje grananje lisnih žila. Stanice traheida su mrtve. Imaju izdužen, često vretenast oblik. Duljina im je 1 – 4 mm. Međutim, u golosjemenjačama, na primjer u Araucariji, doseže 10 mm. Stanične stijenke su debele, celulozne i često impregnirane ligninom. Stanične membrane imaju brojne obrubljene pore.
Žile su nastale u kasnijim fazama evolucije. Karakteristični su za kritosjemenjače, iako ih ima i kod nekih suvremenih predstavnika odjela Mahovine (rod Sellaginella), Preslice, Papratnjače i Golosjemenjače (rod Gnetum).
Žile se sastoje od izduženih mrtvih stanica koje se nalaze jedna iznad druge i nazivaju se segmentima žila. U krajnjim stijenkama segmenata posude nalaze se velike prolazne rupe - perforacije, kroz koje se odvija transport tvari na velike udaljenosti. Perforacije su nastale tijekom evolucije iz obrubljenih pora traheida. As part of the vessels they are ladder and simple. Brojne skalariformne perforacije stvaraju se na krajnjim stijenkama segmenata žile kada su položene koso. Otvori takvih perforacija imaju izduženi oblik, a pregrade koje ih odvajaju nalaze se paralelno jedna s drugom, nalikujući stepenicama stubišta. Posude s skalariformnom perforacijom karakteristične su za biljke iz obitelji Ranunculaceae, Limonaceae, Birch, Palm i Chastukhova.
Jednostavne perforacije poznate su u evolucijski mlađim obiteljima, kao što su Solanaceae, Cucurbitaceae, Asteraceae i Poaceae. Oni predstavljaju jednu veliku rupu u krajnjoj stijenci zgloba, koja se nalazi okomito na os posude. U brojnim obiteljima, na primjer, Magnoliaceae, Roseaceae, Irisaceae, Asteraceae, u posudama se nalaze i jednostavne i skalariformne perforacije.
Bočne stijenke imaju nejednaka celulozna zadebljanja koja štite krvne žile od prekomjernog pritiska koji stvaraju obližnje žive stanice drugih tkiva. Na bočnim stijenkama mogu postojati brojne pore koje omogućuju izlazak vode izvan posude.
Ovisno o prirodi zadebljanja, vrsti i prirodi položaja pora, posude se dijele na prstenaste, spiralne, bispiralne, retikularne, skalariformne i točkaste pore. U prstenastim i spiralnim posudama celulozna zadebljanja raspoređena su u obliku prstenova ili spirala. Kroz nezadebljana područja transportirane otopine difundiraju u okolna tkiva. Promjer ovih posuda je relativno mali. U mrežastim, skalariformnim i točkasto-porastim posudama cijela je bočna stijenka, s izuzetkom mjesta jednostavnih pora, zadebljana i često impregnirana ligninom. Stoga se njihov radijalni transport tvari odvija kroz brojne izdužene i oštre pore.
Plovila imaju ograničen vijek trajanja. One mogu biti uništene kao posljedica začepljenja tilima - izraslinama susjednih stanica parenhima, kao i pod utjecajem sila centripetalnog pritiska novih stanica drva koje stvara kambij. Tijekom evolucije krvne žile prolaze kroz promjene. Segmenti žila postaju kraći i deblji, kose poprečne pregrade zamjenjuju se ravnima, a skalariformne perforacije postaju jednostavne.
Provodna tkiva silazne struje
Silazna tkiva uključuju sitaste stanice i sitaste cijevi sa pratećim stanicama. Sitaste stanice imaju starije podrijetlo. Nalaze se u višim spornim biljkama i golosjemenjačama. To su žive, izdužene stanice sa šiljastim krajevima. U zrelom stanju sadrže jezgre kao dio protoplasta. U njihovim bočnim stijenkama, na mjestima kontakta susjednih stanica, postoje male prolazne perforacije, koje se skupljaju u skupine i tvore sitasta polja kroz koja se kreću tvari.
Sitaste cijevi sastoje se od okomitog niza izduženih ćelija međusobno odvojenih poprečnim stijenkama koje se nazivaju sitaste ploče, u kojima se nalaze sitasta polja. Ako sitasta ploča ima jedno polje sita, smatra se jednostavnom, a ako ih ima više, smatra se složenom. Sitasta polja formiraju brojne prolazne rupe - sitaste perforacije malog promjera. Plazmodezmi prolaze kroz ove perforacije iz jedne stanice u drugu. Na stijenke perforacija nalazi se kalozni polisaharid koji smanjuje lumen perforacija. Kako sitasta cijev stari, kaloza potpuno začepi perforacije i cijev prestaje raditi.
Kada se formira sitasta cijev, u stanicama koje ih tvore sintetizira se poseban protein floema (F-protein) i razvija se velika vakuola. Gura citoplazmu i jezgru prema staničnoj stijenci. Membrana vakuole tada se raspada i unutarnji prostor Stanice su ispunjene mješavinom citoplazme i staničnog soka. Tijela F proteina gube svoje jasne obrise i stapaju se, tvoreći niti u blizini sitastih ploča. Njihova vlakna prolaze kroz perforacije iz jednog segmenta sitaste cijevi u drugi. Jedna ili dvije prateće stanice, koje imaju izdužen oblik, tanke stijenke i živu citoplazmu s jezgrom i brojnim mitohondrijima, tijesno su uz segmente sitaste cijevi. Mitohondriji sintetiziraju ATP, koji je neophodan za prijenos tvari kroz sitaste cijevi. U stijenkama stanica pratilica nalazi se veliki broj pore s plazmadezmama, što je gotovo 10 puta više od njihovog broja u ostalim mezofilnim stanicama lista. Površina protoplasta ovih stanica značajno je povećana zbog brojnih nabora koje stvara plazmalema.
Brzina kretanja asimilata kroz sitaste cijevi znatno premašuje brzinu slobodne difuzije tvari i doseže 50-150 cm / sat, što ukazuje na aktivni transport tvari korištenjem energije ATP.
Trajanje rada sitastih cijevi u višegodišnjim dvosupnicama je 1-2 godine. Da bi ih zamijenio, kambij stalno stvara nove vodljive elemente. Kod jednosupnica koje nemaju kambij, sitaste cijevi traju puno dulje.
Vodljivi snopovi
Provodna tkiva nalaze se u biljnim organima u obliku uzdužnih vrpci, tvoreći vodljive snopove. Postoje četiri vrste vaskularnih snopova: jednostavni, opći, složeni i fibrovaskularni.
Jednostavni snopovi sastoje se od jedne vrste vodljivog tkiva. Na primjer, u rubnim dijelovima lisnih ploča mnogih biljaka nalaze se snopovi posuda i traheida malog promjera, au cvjetnim izdancima ljiljana - samo od sitastih cijevi.
Zajedničke snopove tvore traheide, žile i sitaste cijevi. Ponekad se izraz koristi za označavanje snopova metamera koji prolaze kroz internodije i predstavljaju lisne staze. Složeni snopovi uključuju vodljiva i parenhimska tkiva. Najnapredniji, raznoliki u strukturi i položaju su vaskularno-vlaknasti snopovi.
Vaskularno-vlaknasti snopovi karakteristični su za mnoge više spore biljke i golosjemenjače. Međutim, oni su najtipičniji za angiosperme. U takvim snopovima razlikuju se funkcionalno različiti dijelovi - floem i ksilem. Floem osigurava otjecanje asimilata iz lista i njihovo kretanje do mjesta uporabe ili skladištenja. Ksilem prenosi vodu i u njoj otopljene tvari od korijenskog sustava do lista i drugih organa. Volumen dijela ksilema nekoliko je puta veći od volumena floemskog dijela, budući da volumen vode koja ulazi u biljku premašuje volumen nastalih asimilata, budući da biljka isparava značajan dio vode.
Raznolikost vaskularno-vlaknastih snopova određena je njihovim podrijetlom, histološkim sastavom i smještajem u biljci. Ako se snopovi formiraju iz prokambija i dovrše svoj razvoj jer se koristi opskrba stanica obrazovnog tkiva, kao u jednosupnicama, nazivaju se zatvorenima za rast. Nasuprot tome, kod dikotiledona otvoreni čuperci nisu ograničeni u rastu, budući da ih formira kambij i povećavaju promjer tijekom života biljke. Osim provodnih tkiva, vaskularno-vlaknasti snopovi mogu uključivati osnovna i mehanička tkiva. Na primjer, kod dvosupnica, floem se sastoji od sitastih cijevi (uzlazno tkivo), ličnog parenhima (osnovno tkivo) i ličnih vlakana (mehaničko tkivo). Ksilem se sastoji od žila i traheida (vodljivo tkivo silazne struje), drvnog parenhima (osnovno tkivo) i drvenih vlakana (mehaničko tkivo). Histološki sastav ksilema i floema genetski je određen i može se koristiti u taksonomiji biljaka za dijagnosticiranje različitih svojti. Osim toga, stupanj razvoja komponente grozdovi se mogu mijenjati pod utjecajem uvjeta rasta biljaka.
Poznato je nekoliko vrsta vaskularno-fibroznih snopova.
Zatvoreni kolateralni vaskularni snopovi karakteristični su za listove i stabljike jednosupnica angiospermi. Nedostaje im kambij. Floem i ksilem nalaze se jedan pored drugog. Karakteriziraju ih neki značajke dizajna. Tako se kod pšenice, koja se razlikuje po C 3 -putu fotosinteze, snopovi formiraju iz prokambija i imaju primarni floem i primarni ksilem. U floemu postoji raniji protofloem i kasniji, ali veći metafloem stanica. U floemskom dijelu nedostaje likasti parenhim i likasta vlakna. U ksilemu se u početku stvaraju manje protoksilemske žile, smještene u jednoj liniji okomito na unutarnju granicu floema. Metaksilem je predstavljen s dvije velike posude smještene uz metafloem okomito na lanac protoksilemskih posuda. U ovom slučaju, posude su raspoređene u obliku slova T. Poznat je i raspored posuda u obliku slova V, Y i È. Između metaksilemskih žila, u 1-2 reda, nalazi se sitnostanični sklerenhim zadebljalih stijenki, koji se razvojem stabljike zasiti ligninom. Ovaj sklerenhim odvaja zonu ksilema od floema. S obje strane protoksilemskih žila nalaze se stanice drvenog parenhima, koje vjerojatno igraju transfuzijsku ulogu, budući da tijekom prijelaza snopa iz internodija u jastučić lista čvora stabljike sudjeluju u stvaranju prijenosnih stanica. Oko žilnog snopa stabljike pšenice nalazi se sklerenhimska ovojnica, bolje razvijena na strani protoksilema i protofloema; u blizini bočnih strana snopa stanice ovojnice su raspoređene u jednom redu.
Kod biljaka s C 4 tipom fotosinteze (kukuruz, proso i dr.), u lišću oko zatvorenih vaskularnih snopova nalazi se obloga od velikih stanica klorenhima.

Otvoreni kolateralni snopovi karakteristični su za stabljike dikotiledona. Prisutnost sloja kambija između floema i ksilema, kao i nepostojanje sklerenhimskog omotača oko snopova, osigurava njihov dugotrajni rast u debljinu. U dijelovima ksilema i floema takvih snopova nalaze se stanice glavnog i mehaničkog tkiva.
Otvoreni kolateralni paketi mogu se formirati na dva načina. Prvo, to su snopovi koje primarno formira prokambij. Zatim se iz stanica glavnog parenhima u njima razvija kambij koji proizvodi sekundarne elemente floema i ksilema. Kao rezultat toga, snopovi će kombinirati histološke elemente primarnog i sekundarnog podrijetla. Takvi grozdovi karakteristični su za mnoge zeljaste cvjetnice iz razreda dikotiledona, koje imaju grozdastu strukturu stabljike (mahunarke, Rosaceae itd.).
Drugo, otvorene kolateralne snopove može formirati samo kambij i sastoje se od ksilema i floema sekundarnog podrijetla. Tipični su za zeljaste dikotiledone s prijelaznim tipom anatomske strukture stabljike (asteraceae, itd.), Kao i za korijenske usjeve poput repe.
U stabljikama biljaka brojnih obitelji (Bundeva, Solanaceae, Campanaceae itd.) Postoje otvoreni bikolateralni snopovi, gdje je ksilem s obje strane okružen floemom. U ovom slučaju, vanjski dio floema, okrenut prema površini stabljike, bolje je razvijen od unutarnjeg, a traka kambija se u pravilu nalazi između ksilema i vanjskog dijela floema.
Koncentrične grede dolaze u dvije vrste. Kod amfikibralnih snopova, karakterističnih za rizome paprati, floem okružuje ksilem; kod amfivazalnih snopova ksilem se nalazi u prstenu oko floema (rizomi perunike, đurđice i dr.). Koncentrični snopići rjeđi su kod dvosupnica (ricinusa).
Zatvoreni radijalni vaskularni snopovi formiraju se u područjima korijena koji imaju primarni anatomska građa. Radijalni snop dio je središnjeg cilindra i prolazi kroz sredinu korijena. Njegov ksilem ima izgled višezračne zvijezde. Stanice floema nalaze se između ksilemskih zraka. Broj ksilemskih zraka uvelike ovisi o genetskoj prirodi biljaka. Na primjer, kod mrkve, repe, kupusa i drugih dikotiledona, ksilem radijalnog snopa ima samo dvije zrake. Stabla jabuke i kruške mogu ih imati 3-5, bundeve i grah imaju četveroprugasti ksilem, a jednosupnice imaju višezrakasti ksilem. Radijalni raspored ksilemskih zraka ima adaptivno značenje. Skraćuje put vode od usisne površine korijena do žila središnjeg cilindra.
Kod višegodišnjih drvenastih biljaka i nekih zeljastih jednogodišnjih biljaka, kao što je lan, vaskularna tkiva nalaze se u stabljici bez formiranja jasno definiranih vaskularnih snopova. Zatim govore o nečupavom tipu strukture stabljike.
Tkiva koja reguliraju radijalni transport tvari
Specifična tkiva koja reguliraju radijalni transport tvari uključuju egzoderm i endoderm.

Egzoderm je vanjski sloj kore primarnog korijena. Nastaje neposredno ispod primarnog integumentarnog epiblemskog tkiva u zoni korijenovih dlačica i sastoji se od jednog ili više slojeva tijesno zbijenih stanica sa zadebljalim celuloznim membranama. U egzodermisu voda koja ulazi u korijen kroz korijenove dlačice doživljava otpor viskozne citoplazme i prelazi u celulozne membrane egzodermalnih stanica, a zatim ih napušta u međustaničnim prostorima srednjeg sloja primarnog korteksa ili mezoderma. Time se osigurava učinkovit protok vode u dublje slojeve korijena.
U zoni provođenja u korijenu jednosupnica, gdje stanice epibleme odumiru i ljušte se, na površini korijena pojavljuje se egzodermis. Njegove stanične stijenke zasićene su suberinom i sprječavaju protok vode iz tla u korijen. Kod dikotiledona, egzoderm u primarnom korteksu se skida tijekom linjanja korijena i zamjenjuje peridermom.
Endoderm ili unutarnji sloj kore primarnog korijena nalazi se oko središnjeg cilindra. Tvori ga jedan sloj čvrsto zatvorenih stanica nejednake strukture. Neki od njih, koji se nazivaju propusni, imaju tanke ljuske i lako propuštaju vodu. Kroz njih voda iz primarne kore ulazi u radijalni vaskularni snop korijena. Ostale stanice imaju specifična celulozna zadebljanja radijalnih i unutarnjih tangencijalnih stijenki. Ova zadebljanja, zasićena suberinom, nazivaju se kasparski pojasevi. Nepropusni su za vodu. Stoga voda ulazi u središnji cilindar samo kroz prolazne ćelije. A budući da upijajuća površina korijena značajno premašuje ukupnu površinu poprečnog presjeka stanica endodermalnog prolaza, nastaje pritisak korijena, što je jedan od mehanizama protoka vode u stabljiku, list i reproduktivne organe.
Endoderm je također dio kore mlade stabljike. U nekim zeljastim angiospermama, kao i korijen, može imati kasparske pojaseve. Osim toga, kod mladih stabljika endodermis može biti predstavljen omotačem koji nosi škrob. Na taj način endodermis može regulirati transport vode u biljci i skladištiti hranjive tvari.
Pojam stele i njezina evolucija
Mnogo se pozornosti posvećuje nastanku, razvoju u ontogenezi i evolucijskim strukturnim preobrazbama provodnog sustava, budući da osigurava međusobnu povezanost biljnih organa i s njim je povezana evolucija velikih svojti.
Na prijedlog francuskih botaničara F. Van Tieghema i A. Dulia (1886.), skup primarnih provodnih tkiva, zajedno s drugim tkivima koji se nalaze između njih i pericikla uz koru, nazvan je stela. Stela također može uključivati jezgru i šupljinu formiranu na njenom mjestu, kao, na primjer, u bluegrassu. Koncept "stele" odgovara konceptu "centralnog cilindra". Stela korijena i stabljike funkcionalno je objedinjena. Proučavanje stele kod predstavnika različitih odjela viših biljaka dovelo je do formiranja teorije stele.
Postoje dvije glavne vrste stela: protostele i eustele. Najstariji je protostela. Njegova provodna tkiva nalaze se u sredini aksijalnih organa, sa ksilemom u središtu, okruženim kontinuiranim slojem floema. U stabljici nema srži niti šupljine.
Postoji nekoliko evolucijski povezanih tipova protostela: haplostele, aktinostele i plektostele.
Izvorni, primitivni tip je haplostela. Njegov ksilem ima zaobljen presjek i okružen je ravnomjernim, kontinuiranim slojem floema. Pericikl se nalazi oko provodnih tkiva u jednom ili dva sloja. Haplostela je bila poznata iz fosilnih riniofita i sačuvana je u nekim psilotofitima (tmesipterus).
Više razvijene vrste Protostela je aktinostela kod koje ksilem u presjeku ima oblik višezrakaste zvijezde. Nalazi se u fosilima Asteroxylona i nekim primitivnim likofitima.
Daljnje odvajanje ksilema u odvojene dijelove smještene radijalno ili paralelno jedan s drugim dovelo je do stvaranja plektostele, karakteristične za stabljike likofita. Kod aktinostele i plektostele floem još uvijek okružuje ksilem sa svih strana.
Tijekom evolucije sifonostela je nastala iz protostele, razlikovna značajka koji je cjevaste strukture. U središtu takve stele nalazi se jezgra ili šupljina. U provodnom dijelu sifonostele pojavljuju se lisni prorezi, zahvaljujući kojima dolazi do kontinuirane veze između jezgre i kore. Ovisno o načinu međusobnog rasporeda ksilema i floema, sifonostela je ektofloična i amfifloična. U prvom slučaju floem okružuje ksilem s jedne, vanjske strane. U drugom, floem okružuje ksilem s obje strane, vanjske i unutarnje.
Kada se amfiflojski sifonostel podijeli u mrežu ili nizove uzdužnih niti, pojavljuje se rascjepkana stela ili diktiostela, karakteristična za mnoge papratnjače. Njegov provodni dio predstavljen je brojnim koncentričnim provodnim snopovima.
Kod preslica je artrostela nastala iz ektofloičnog sifonostela koji ima segmentiranu strukturu. Razlikuje se prisutnošću jedne velike središnje šupljine i zasebnih vaskularnih snopova s protoksilemskim šupljinama (karinalni kanali).
Kod cvjetnica, na temelju ektofloičnog sifonostela, formirana je eustela, karakteristična za dikotiledone, i atakostela, tipična za jednosupnice. U eusteli, provodni dio sastoji se od zasebnih kolateralnih snopova koji imaju kružni raspored. U središtu stele u stabljici nalazi se jezgra, koja je uz pomoć medularnih zraka povezana s korom. U ataxosteleu, vaskularni snopovi imaju raštrkan raspored, između njih nalaze se stanice parenhima središnjeg cilindra. Ovakav raspored greda skriva cjevasta struktura sifonostele.
Pojava razne opcije siphonosteles su važna prilagodba viših biljaka za povećanje promjera aksijalnih organa - korijena i stabljike.
6.1. Značaj i raznolikost provodnih tkiva
Provodna tkiva najvažniji su sastavni dio većine viših biljaka. Obvezna su strukturna komponenta vegetativnih i rasplodnih organa spornih i sjemenskih biljaka. Provodna tkiva zajedno sa staničnom stjenkom i međustaničnim prostorima, nekim stanicama glavnog parenhima i specijaliziranim prijenosnim stanicama čine provodni sustav koji osigurava daljinski i radijalni transport tvari. Zbog posebnog dizajna stanica i njihovog položaja u tijelu biljke, provodni sustav obavlja brojne, ali međusobno povezane funkcije:
1) kretanje vode i minerala koje korijenje apsorbira iz tla, kao i organskih tvari nastalih u korijenju, u stabljiku, lišće i reproduktivne organe;
2) kretanje produkata fotosinteze od zelenih dijelova biljke do mjesta njihove upotrebe i skladištenja: korijenje, stabljike, plodovi i sjemenke;
3) kretanje fitohormona kroz biljku, čime se stvara određena njihova ravnoteža, koja određuje brzinu rasta i razvoja vegetativnih i reproduktivnih organa biljaka;
4) radijalni transport tvari iz provodnih tkiva u obližnje žive stanice drugih tkiva, na primjer, u asimilirajuće mezofilne stanice listova i diobene stanice meristema. U njemu mogu sudjelovati i parenhimske stanice medularnih zraka drva i kore. Transmisivne stanice s brojnim izbočinama stanične membrane, smještene između provodnog i parenhimskog tkiva, imaju veliku važnost u radijalnom transportu;
5) vodljiva tkiva povećavaju otpornost biljnih organa na deformirajuća opterećenja;
6) provodna tkiva čine kontinuirani razgranati sustav koji povezuje biljne organe u jedinstvenu cjelinu;
Pojava provodnih tkiva rezultat je evolucijskih strukturnih transformacija povezanih s izlaskom biljaka na kopno i odvajanjem njihove prehrane iz zraka i tla. Najstarija provodna tkiva, traheide, pronađena su u fosilnim rinofitima. Svoj najveći razvoj dosegle su u modernim angiospermama.
Tijekom procesa individualnog razvoja iz prokambija nastaju primarna provodna tkiva na mjestima rasta sjemenog zametka i pupoljaka obnove. Sekundarna provodna tkiva, karakteristična za dikotiledone angiosperme, stvara kambij.
Ovisno o funkcijama koje obavljaju, provodna tkiva se dijele na tkiva uzlazne struje i tkiva silazne struje. Glavna svrha uzlaznog tkiva je prijenos vode i u njoj otopljenih minerala od korijena do viših nadzemnih organa. Osim toga, kroz njih se kreću organske tvari nastale u korijenu i stabljici, kao što su organske kiseline, ugljikohidrati i fitohormoni. Međutim, izraz "uzlazna struja" ne treba jednoznačno shvatiti kao kretanje odozdo prema gore. Uzlazna tkiva osiguravaju protok tvari u smjeru od usisne zone do vrha izdanka. U tom slučaju transportirane tvari koristi kako sam korijen, tako i stabljika, grane, lišće, reproduktivni organi, bez obzira nalaze li se iznad ili ispod razine korijena. Na primjer, kod krumpira voda i elementi mineralne prehrane teku kroz uzlazna tkiva u stolone i gomolje formirane u tlu, kao iu nadzemne organe.
Nizvodna tkiva osiguravaju odljev produkata fotosinteze u rastuće dijelove biljaka i organe za pohranu. U ovom slučaju prostorni položaj fotosintetskih organa nije bitan. Na primjer, u pšenici, organske tvari ulaze u razvoj zrna iz listova različitih slojeva. Stoga nazive "uzlaznih" i "silaznih" tkanina treba tretirati kao ništa više od utvrđene tradicije.
6.2. Provodna tkiva uzlazne struje
U uzlazna tkiva spadaju traheide i žile (traheje), koje se nalaze u drvenastom (ksilemskom) dijelu biljnih organa. U tim tkivima kretanje vode i u njoj otopljenih tvari odvija se pasivno pod utjecajem pritiska korijena i isparavanja vode s površine biljke.
Traheidi su starijeg porijekla. Nalaze se u višim spornim biljkama, golosjemenjačama, a rjeđe u kritosjemenjačama. Kod angiospermi su tipične za najmanje grananje lisnih žila. Stanice traheida su mrtve. Imaju izdužen, često vretenast oblik. Duljina im je 1 – 4 mm. Međutim, u golosjemenjačama, na primjer u Araucariji, doseže 10 mm. Stanične stijenke su debele, celulozne i često impregnirane ligninom. Stanične membrane imaju brojne obrubljene pore.
Žile su nastale u kasnijim fazama evolucije. Karakteristični su za kritosjemenjače, iako ih ima i kod nekih suvremenih predstavnika odjela Mahovine (rod Sellaginella), Preslice, Papratnjače i Golosjemenjače (rod Gnetum).
Žile se sastoje od izduženih mrtvih stanica koje se nalaze jedna iznad druge i nazivaju se segmentima žila. U krajnjim stijenkama segmenata posude nalaze se velike prolazne rupe - perforacije, kroz koje se odvija transport tvari na velike udaljenosti. Perforacije su nastale tijekom evolucije iz obrubljenih pora traheida. As part of the vessels they are ladder and simple. Brojne skalariformne perforacije stvaraju se na krajnjim stijenkama segmenata žile kada su položene koso. Otvori takvih perforacija imaju izduženi oblik, a pregrade koje ih odvajaju nalaze se paralelno jedna s drugom, nalikujući stepenicama stubišta. Posude s skalariformnom perforacijom karakteristične su za biljke iz obitelji Ranunculaceae, Limonaceae, Birch, Palm i Chastukhova.
Jednostavne perforacije poznate su u evolucijski mlađim obiteljima, kao što su Solanaceae, Cucurbitaceae, Asteraceae i Poaceae. Oni predstavljaju jednu veliku rupu u krajnjoj stijenci zgloba, koja se nalazi okomito na os posude. U brojnim obiteljima, na primjer, Magnoliaceae, Roseaceae, Irisaceae, Asteraceae, u posudama se nalaze i jednostavne i skalariformne perforacije.
Bočne stijenke imaju nejednaka celulozna zadebljanja koja štite krvne žile od prekomjernog pritiska koji stvaraju obližnje žive stanice drugih tkiva. Na bočnim stijenkama mogu postojati brojne pore koje omogućuju izlazak vode izvan posude.
Ovisno o prirodi zadebljanja, vrsti i prirodi položaja pora, posude se dijele na prstenaste, spiralne, bispiralne, retikularne, skalariformne i točkaste pore. U prstenastim i spiralnim posudama celulozna zadebljanja raspoređena su u obliku prstenova ili spirala. Kroz nezadebljana područja transportirane otopine difundiraju u okolna tkiva. Promjer ovih posuda je relativno mali. U mrežastim, skalariformnim i točkasto-porastim posudama cijela je bočna stijenka, s izuzetkom mjesta jednostavnih pora, zadebljana i često impregnirana ligninom. Stoga se njihov radijalni transport tvari odvija kroz brojne izdužene i oštre pore.
Plovila imaju ograničen vijek trajanja. One mogu biti uništene kao posljedica začepljenja tilima - izraslinama susjednih stanica parenhima, kao i pod utjecajem sila centripetalnog pritiska novih stanica drva koje stvara kambij. Tijekom evolucije krvne žile prolaze kroz promjene. Segmenti žila postaju kraći i deblji, kose poprečne pregrade zamjenjuju se ravnima, a skalariformne perforacije postaju jednostavne.
6.3. Provodna tkiva silazne struje
Silazna tkiva uključuju sitaste stanice i sitaste cijevi sa pratećim stanicama. Sitaste stanice imaju starije podrijetlo. Nalaze se u višim spornim biljkama i golosjemenjačama. To su žive, izdužene stanice sa šiljastim krajevima. U zrelom stanju sadrže jezgre kao dio protoplasta. U njihovim bočnim stijenkama, na mjestima kontakta susjednih stanica, postoje male prolazne perforacije, koje se skupljaju u skupine i tvore sitasta polja kroz koja se kreću tvari.
Sitaste cijevi sastoje se od okomitog niza izduženih ćelija međusobno odvojenih poprečnim stijenkama koje se nazivaju sitaste ploče, u kojima se nalaze sitasta polja. Ako sitasta ploča ima jedno polje sita, smatra se jednostavnom, a ako ih ima više, smatra se složenom. Sitasta polja formiraju brojne prolazne rupe - sitaste perforacije malog promjera. Plazmodezmi prolaze kroz ove perforacije iz jedne stanice u drugu. Na stijenke perforacija nalazi se kalozni polisaharid koji smanjuje lumen perforacija. Kako sitasta cijev stari, kaloza potpuno začepi perforacije i cijev prestaje raditi.
Kada se formira sitasta cijev, u stanicama koje ih tvore sintetizira se poseban protein floema (F-protein) i razvija se velika vakuola. Gura citoplazmu i jezgru prema staničnoj stijenci. Tada dolazi do razaranja membrane vakuole, a unutarnji prostor stanice ispunjen je mješavinom citoplazme i staničnog soka. Tijela F proteina gube svoje jasne obrise i stapaju se, tvoreći niti u blizini sitastih ploča. Njihova vlakna prolaze kroz perforacije iz jednog segmenta sitaste cijevi u drugi. Jedna ili dvije prateće stanice, koje imaju izdužen oblik, tanke stijenke i živu citoplazmu s jezgrom i brojnim mitohondrijima, tijesno su uz segmente sitaste cijevi. Mitohondriji sintetiziraju ATP, koji je neophodan za prijenos tvari kroz sitaste cijevi. U stijenkama stanica pratilica nalazi se veliki broj pora s plazmadezmatima, koji je gotovo 10 puta veći od njihovog broja u ostalim mezofilnim stanicama lista. Površina protoplasta ovih stanica značajno je povećana zbog brojnih nabora koje stvara plazmalema.
Brzina kretanja asimilata kroz sitaste cijevi znatno premašuje brzinu slobodne difuzije tvari i doseže 50-150 cm / sat, što ukazuje na aktivni transport tvari korištenjem energije ATP.
Trajanje rada sitastih cijevi u višegodišnjim dvosupnicama je 1-2 godine. Da bi ih zamijenio, kambij stalno stvara nove vodljive elemente. Kod jednosupnica koje nemaju kambij, sitaste cijevi traju puno dulje.
6.4. Vodljivi snopovi
Provodna tkiva nalaze se u biljnim organima u obliku uzdužnih vrpci, tvoreći vodljive snopove. Postoje četiri vrste vaskularnih snopova: jednostavni, opći, složeni i fibrovaskularni.
Jednostavni snopovi sastoje se od jedne vrste vodljivog tkiva. Na primjer, u rubnim dijelovima lisnih ploča mnogih biljaka nalaze se snopovi posuda i traheida malog promjera, au cvjetnim izdancima ljiljana - samo od sitastih cijevi.
Zajedničke snopove tvore traheide, žile i sitaste cijevi. Ponekad se izraz koristi za označavanje snopova metamera koji prolaze kroz internodije i predstavljaju lisne staze. Složeni snopovi uključuju vodljiva i parenhimska tkiva. Najnapredniji, raznoliki u strukturi i položaju su vaskularno-vlaknasti snopovi.
Vaskularno-vlaknasti snopovi karakteristični su za mnoge više spore biljke i golosjemenjače. Međutim, oni su najtipičniji za angiosperme. U takvim snopovima razlikuju se funkcionalno različiti dijelovi - floem i ksilem. Floem osigurava otjecanje asimilata iz lista i njihovo kretanje do mjesta uporabe ili skladištenja. Ksilem prenosi vodu i u njoj otopljene tvari od korijenskog sustava do lista i drugih organa. Volumen dijela ksilema nekoliko je puta veći od volumena floemskog dijela, budući da volumen vode koja ulazi u biljku premašuje volumen nastalih asimilata, budući da biljka isparava značajan dio vode.
Raznolikost vaskularno-vlaknastih snopova određena je njihovim podrijetlom, histološkim sastavom i smještajem u biljci. Ako se snopovi formiraju iz prokambija i dovrše svoj razvoj jer se koristi opskrba stanica obrazovnog tkiva, kao u jednosupnicama, nazivaju se zatvorenima za rast. Nasuprot tome, kod dikotiledona otvoreni čuperci nisu ograničeni u rastu, budući da ih formira kambij i povećavaju promjer tijekom života biljke. Osim provodnih tkiva, vaskularno-vlaknasti snopovi mogu uključivati osnovna i mehanička tkiva. Na primjer, kod dvosupnica, floem se sastoji od sitastih cijevi (uzlazno tkivo), ličnog parenhima (osnovno tkivo) i ličnih vlakana (mehaničko tkivo). Ksilem se sastoji od žila i traheida (vodljivo tkivo silazne struje), drvnog parenhima (osnovno tkivo) i drvenih vlakana (mehaničko tkivo). Histološki sastav ksilema i floema genetski je određen i može se koristiti u taksonomiji biljaka za dijagnosticiranje različitih svojti. Osim toga, stupanj razvoja sastavnih dijelova grozdova može se promijeniti pod utjecajem uvjeta rasta biljaka.
Poznato je nekoliko vrsta vaskularno-fibroznih snopova.
Zatvoreni kolateralni vaskularni snopovi karakteristični su za listove i stabljike jednosupnica angiospermi. Nedostaje im kambij. Floem i ksilem nalaze se jedan pored drugog. Karakteriziraju ih određene značajke dizajna. Tako se kod pšenice, koja se razlikuje po C 3 -putu fotosinteze, snopovi formiraju iz prokambija i imaju primarni floem i primarni ksilem. U floemu postoji raniji protofloem i kasniji, ali veći metafloem stanica. U floemskom dijelu nedostaje likasti parenhim i likasta vlakna. U ksilemu se u početku stvaraju manje protoksilemske žile, smještene u jednoj liniji okomito na unutarnju granicu floema. Metaksilem je predstavljen s dvije velike posude smještene uz metafloem okomito na lanac protoksilemskih posuda. U ovom slučaju, posude su raspoređene u obliku slova T. Poznat je i raspored posuda u obliku slova V, Y i È. Između metaksilemskih žila, u 1-2 reda, nalazi se sitnostanični sklerenhim zadebljalih stijenki, koji se razvojem stabljike zasiti ligninom. Ovaj sklerenhim odvaja zonu ksilema od floema. S obje strane protoksilemskih žila nalaze se stanice drvenog parenhima, koje vjerojatno igraju transfuzijsku ulogu, budući da tijekom prijelaza snopa iz internodija u jastučić lista čvora stabljike sudjeluju u stvaranju prijenosnih stanica. Oko žilnog snopa stabljike pšenice nalazi se sklerenhimska ovojnica, bolje razvijena na strani protoksilema i protofloema; u blizini bočnih strana snopa stanice ovojnice su raspoređene u jednom redu.
Kod biljaka s C 4 tipom fotosinteze (kukuruz, proso i dr.), u lišću oko zatvorenih vaskularnih snopova nalazi se obloga od velikih stanica klorenhima.
Otvoreni kolateralni snopovi karakteristični su za stabljike dikotiledona. Prisutnost sloja kambija između floema i ksilema, kao i nepostojanje sklerenhimskog omotača oko snopova, osigurava njihov dugotrajni rast u debljinu. U dijelovima ksilema i floema takvih snopova nalaze se stanice glavnog i mehaničkog tkiva.
Otvoreni kolateralni paketi mogu se formirati na dva načina. Prvo, to su snopovi koje primarno formira prokambij. Zatim se iz stanica glavnog parenhima u njima razvija kambij koji proizvodi sekundarne elemente floema i ksilema. Kao rezultat toga, snopovi će kombinirati histološke elemente primarnog i sekundarnog podrijetla. Takvi grozdovi karakteristični su za mnoge zeljaste cvjetnice iz razreda dikotiledona, koje imaju grozdastu strukturu stabljike (mahunarke, Rosaceae itd.).
Drugo, otvorene kolateralne snopove može formirati samo kambij i sastoje se od ksilema i floema sekundarnog podrijetla. Tipični su za zeljaste dikotiledone s prijelaznim tipom anatomske strukture stabljike (asteraceae, itd.), Kao i za korijenske usjeve poput repe.
U stabljikama biljaka brojnih obitelji (Bundeva, Solanaceae, Campanaceae itd.) Postoje otvoreni bikolateralni snopovi, gdje je ksilem s obje strane okružen floemom. U ovom slučaju, vanjski dio floema, okrenut prema površini stabljike, bolje je razvijen od unutarnjeg, a traka kambija se u pravilu nalazi između ksilema i vanjskog dijela floema.
Koncentrične grede dolaze u dvije vrste. Kod amfikibralnih snopova, karakterističnih za rizome paprati, floem okružuje ksilem; kod amfivazalnih snopova ksilem se nalazi u prstenu oko floema (rizomi perunike, đurđice i dr.). Koncentrični snopići rjeđi su kod dvosupnica (ricinusa).
Zatvoreni radijalni vaskularni snopovi formiraju se u područjima korijena koji imaju primarnu anatomsku strukturu. Radijalni snop dio je središnjeg cilindra i prolazi kroz sredinu korijena. Njegov ksilem ima izgled višezračne zvijezde. Stanice floema nalaze se između ksilemskih zraka. Broj ksilemskih zraka uvelike ovisi o genetskoj prirodi biljaka. Na primjer, kod mrkve, repe, kupusa i drugih dikotiledona, ksilem radijalnog snopa ima samo dvije zrake. Stabla jabuke i kruške mogu ih imati 3-5, bundeve i grah imaju četveroprugasti ksilem, a jednosupnice imaju višezrakasti ksilem. Radijalni raspored ksilemskih zraka ima adaptivno značenje. Skraćuje put vode od usisne površine korijena do žila središnjeg cilindra.
Kod višegodišnjih drvenastih biljaka i nekih zeljastih jednogodišnjih biljaka, kao što je lan, vaskularna tkiva nalaze se u stabljici bez formiranja jasno definiranih vaskularnih snopova. Zatim govore o nečupavom tipu strukture stabljike.
6.5. Tkiva koja reguliraju radijalni transport tvari
Specifična tkiva koja reguliraju radijalni transport tvari uključuju egzoderm i endoderm.
Egzoderm je vanjski sloj kore primarnog korijena. Nastaje neposredno ispod primarnog integumentarnog epiblemskog tkiva u zoni korijenovih dlačica i sastoji se od jednog ili više slojeva tijesno zbijenih stanica sa zadebljalim celuloznim membranama. U egzodermisu voda koja ulazi u korijen kroz korijenove dlačice doživljava otpor viskozne citoplazme i prelazi u celulozne membrane egzodermalnih stanica, a zatim ih napušta u međustaničnim prostorima srednjeg sloja primarnog korteksa ili mezoderma. Time se osigurava učinkovit protok vode u dublje slojeve korijena.
U zoni provođenja u korijenu jednosupnica, gdje stanice epibleme odumiru i ljušte se, na površini korijena pojavljuje se egzodermis. Njegove stanične stijenke zasićene su suberinom i sprječavaju protok vode iz tla u korijen. Kod dikotiledona, egzoderm u primarnom korteksu se skida tijekom linjanja korijena i zamjenjuje peridermom.
Endoderm ili unutarnji sloj kore primarnog korijena nalazi se oko središnjeg cilindra. Tvori ga jedan sloj čvrsto zatvorenih stanica nejednake strukture. Neki od njih, koji se nazivaju propusni, imaju tanke ljuske i lako propuštaju vodu. Kroz njih voda iz primarne kore ulazi u radijalni vaskularni snop korijena. Ostale stanice imaju specifična celulozna zadebljanja radijalnih i unutarnjih tangencijalnih stijenki. Ova zadebljanja, zasićena suberinom, nazivaju se kasparski pojasevi. Nepropusni su za vodu. Stoga voda ulazi u središnji cilindar samo kroz prolazne ćelije. A budući da upijajuća površina korijena značajno premašuje ukupnu površinu poprečnog presjeka stanica endodermalnog prolaza, nastaje pritisak korijena, što je jedan od mehanizama protoka vode u stabljiku, list i reproduktivne organe.
Endoderm je također dio kore mlade stabljike. U nekim zeljastim angiospermama, kao i korijen, može imati kasparske pojaseve. Osim toga, kod mladih stabljika endodermis može biti predstavljen omotačem koji nosi škrob. Na taj način endodermis može regulirati transport vode u biljci i skladištiti hranjive tvari.
6.6. Pojam stele i njezina evolucija
Mnogo se pozornosti posvećuje nastanku, razvoju u ontogenezi i evolucijskim strukturnim preobrazbama provodnog sustava, budući da osigurava međusobnu povezanost biljnih organa i s njim je povezana evolucija velikih svojti.
Na prijedlog francuskih botaničara F. Van Tieghema i A. Dulia (1886.), skup primarnih provodnih tkiva, zajedno s drugim tkivima koji se nalaze između njih i pericikla uz koru, nazvan je stela. Stela također može uključivati jezgru i šupljinu formiranu na njenom mjestu, kao, na primjer, u bluegrassu. Koncept "stele" odgovara konceptu "centralnog cilindra". Stela korijena i stabljike funkcionalno je objedinjena. Proučavanje stele kod predstavnika različitih odjela viših biljaka dovelo je do formiranja teorije stele.
Postoje dvije glavne vrste stela: protostele i eustele. Najstariji je protostela. Njegova provodna tkiva nalaze se u sredini aksijalnih organa, sa ksilemom u središtu, okruženim kontinuiranim slojem floema. U stabljici nema srži niti šupljine.
Postoji nekoliko evolucijski povezanih tipova protostela: haplostele, aktinostele i plektostele.
Izvorni, primitivni tip je haplostela. Njegov ksilem ima zaobljen presjek i okružen je ravnomjernim, kontinuiranim slojem floema. Pericikl se nalazi oko provodnih tkiva u jednom ili dva sloja [K. Ezav, 1969]. Haplostela je bila poznata iz fosilnih riniofita i sačuvana je u nekim psilotofitima (tmesipterus).
Razvijeniji tip protostele je aktinostela, kod koje ksilem u presjeku ima oblik višezrakaste zvijezde. Nalazi se u fosilima Asteroxylona i nekim primitivnim likofitima.
Daljnje odvajanje ksilema u odvojene dijelove smještene radijalno ili paralelno jedan s drugim dovelo je do stvaranja plektostele, karakteristične za stabljike likofita. Kod aktinostele i plektostele floem još uvijek okružuje ksilem sa svih strana.
Tijekom evolucije iz protostela je nastao sifonostel, čija je posebnost njegova cjevasta struktura. U središtu takve stele nalazi se jezgra ili šupljina. U provodnom dijelu sifonostele pojavljuju se lisni prorezi, zahvaljujući kojima dolazi do kontinuirane veze između jezgre i kore. Ovisno o načinu međusobnog rasporeda ksilema i floema, sifonostela je ektofloična i amfifloična. U prvom slučaju floem okružuje ksilem s jedne, vanjske strane. U drugom, floem okružuje ksilem s obje strane, vanjske i unutarnje.
Kada se amfiflojski sifonostel podijeli u mrežu ili nizove uzdužnih niti, pojavljuje se rascjepkana stela ili diktiostela, karakteristična za mnoge papratnjače. Njegov provodni dio predstavljen je brojnim koncentričnim provodnim snopovima.
Kod preslica je artrostela nastala iz ektofloičnog sifonostela koji ima segmentiranu strukturu. Razlikuje se prisutnošću jedne velike središnje šupljine i zasebnih vaskularnih snopova s protoksilemskim šupljinama (karinalni kanali).
Kod cvjetnica, na temelju ektofloičnog sifonostela, formirana je eustela, karakteristična za dikotiledone, i atakostela, tipična za jednosupnice. U eusteli, provodni dio sastoji se od zasebnih kolateralnih snopova koji imaju kružni raspored. U središtu stele u stabljici nalazi se jezgra, koja je uz pomoć medularnih zraka povezana s korom. U ataxosteleu, vaskularni snopovi imaju raštrkan raspored, između njih nalaze se stanice parenhima središnjeg cilindra. Ovakav raspored snopova skriva cjevastu strukturu sifonostela.
Pojava raznih varijanti sifonostela važna je prilagodba viših biljaka za povećanje promjera aksijalnih organa - korijena i stabljike.
25. 8.1. Ekskretorni sustav biljaka i njegov značaj
Biljni život je genetski određen skup biokemijskih reakcija čija se brzina i intenzitet značajno mijenjaju uvjetima okoliša uzgoja. Ove reakcije proizvode široku paletu nusproizvoda koje biljka ne koristi za izgradnju tijela ili za regulaciju razmjene tvari, energije i informacija s okolinom. Takvi se proizvodi mogu ukloniti iz postrojenja različiti putevi: sa smrću i odvajanjem grana i dijelova rizoma, s opadanjem lišća i ljuštenjem vanjskih slojeva kore, kao rezultat aktivnosti specijaliziranih struktura vanjske i unutarnje sekrecije. Zajedno, ovi uređaji čine sustav izlučivanja biljaka.
Za razliku od životinja, sustav izlučivanja kod biljaka nije usmjeren na uklanjanje dušikovih spojeva, koji se mogu ponovno iskoristiti u procesu života.
Ekskretorni sustav biljaka je višenamjenski. U njegovim strukturama odvijaju se: sinteza, akumulacija, provođenje i otpuštanje produkata metabolizma. Na primjer, u sekretornim stanicama smolnih kanala u lišću crnogoričnog drveća stvara se smola koja se oslobađa kroz smolne kanale. U nektarima cvjetova lipe stvara se i izlučuje slatki nektarski sok. U posebnim spremnicima u voćnoj ljusci agruma nakupljaju se esencijalna ulja.
Stvaranje i otpuštanje metaboličkih nusproizvoda ima različito prilagodbeno značenje:
Privlačenje insekata oprašivača. Cvjetovi jabuke, krastavca i drugih entomofilnih križnih oprašivača proizvode nektar koji privlači pčele, a smrdljive izlučevine cvijeta raflezije privlače muhe;
Odbijanje biljojeda (kumin, kopriva i dr.);
Zaštita od bakterija i gljivica koje uništavaju drvo (bor, smreka i dr.);
Otpuštanje hlapljivih spojeva u atmosferu, što pomaže u pročišćavanju zraka od patogenih bakterija;
Izvanstanična probava plijena kod kukcoždernih biljaka zbog oslobađanja proteolitičkih enzima (rosika, aldrovanda i dr.);
Mineralizacija organskih ostataka u tlu zbog oslobađanja posebnih enzima tla od strane korijena;
Regulacija vodni režim kroz vodene stomate - hidatode smještene duž ruba lisne ploče (jagode, kupus, crassula, itd.);
Regulacija isparavanja vode kao rezultat oslobađanja hlapljivih eterskih spojeva, koji smanjuju prozirnost i toplinsku vodljivost zraka u blizini površine lista (četinari);
Regulacija režima soli stanica (pigweed, quinoa, itd.);
Promjena kemijskih i fizička svojstva tlo, kao i regulacija sastava vrsta mikroflore tla pod utjecajem korijenskih eksudata;
Regulacija međudjelovanja biljaka u fitocenozi putem izlučevina korijena, stabljike i lišća, naziva se alelopatija (luk, češnjak, itd.).
Tvari koje oslobađaju biljke vrlo su raznolike. Njihova priroda ovisi o genotipu biljaka.
Mnoge vrste proizvode vodu (jagode, kupus), soli (svinjska trava, kvinoja), monosaharide i organske kiseline (maslačak, cikorija), nektar (lipa, heljda), aminokiseline i proteine (topola, vrba), eterična ulja (metvica, ruža) ), balzam (jela), smole (bor, smreka), kaučuk (hevea, kok-sagyz), sluzi (stanice korijenove klobuke, bubreće sjemenke raznih biljaka), probavni sokovi (rosika, maslac), otrovne tekućine (kopriva). , hogweed) i druge veze.
26. 1.1. Polaritet
Polarnost je prisutnost biokemijskih, funkcionalnih i strukturnih razlika u dijametralno suprotnim dijelovima organa cijelog biljnog organizma. Polaritet utječe na intenzitet biokemijskih procesa u stanici i funkcionalnu aktivnost organela, te određuje dizajn anatomskih struktura. Na razini cjelovitog biljnog organizma polaritet je povezan sa smjerom rasta i razvoja pod utjecajem gravitacijskih sila.
Fenomen polariteta uočava se na različitim razinama organizacije biljaka. Na molekularnoj razini očituje se u strukturi molekula organskih tvari, prvenstveno nukleinskih kiselina i proteina. Dakle, polarnost lanaca DNA određena je posebnim redoslijedom povezivanja njezinih nukleotida. Polarnost polipeptida povezana je s prisutnošću amino skupine –NH 2 i karboksilne skupine –COOH u sastavu aminokiselina. Polaritet molekula klorofila posljedica je prisutnosti jezgre tetrapirol porfirina i ostataka alkohola - metanola i fitola.
Stanice i njihove organele mogu imati polaritet. Na primjer, na regeneracijskom polu Golgijevog kompleksa nastaju novi diktiosomi, a na sekretornom polu nastaju vezikule koje su povezane s uklanjanjem metaboličkih produkata iz stanice.
Polarnost stanica nastaje tijekom njihove ontogeneze. Polarizacija stanica nastaje kao rezultat pH gradijenata, električni naboji i osmotski potencijal, koncentraciju O 2 i CO 2, katione kalcija, fiziološki aktivne tvari i elemente mineralne prehrane. Može nastati i pod utjecajem mehaničkog pritiska, sila površinske napetosti i utjecajem susjednih stanica.
Polaritet stanica može biti simetričan ili asimetričan. Simetrična polarnost neophodan je uvjet za diobu izvornih stanica i stvaranje ekvivalentnih stanica kćeri. Konkretno, mikrotubule citoskeleta pomiču se u ekvatorijalnu ravninu i sudjeluju u formiranju fragmoplasta; kromosomi se dijele i njihove se kromatide uz pomoć poteznih niti akromatinskog vretena razilaze na suprotne polove stanice. Tu se ribosomi i mitohondriji počinju koncentrirati. Golgijevi kompleksi pomiču se u središte i specifično su orijentirani u prostoru. Njihovi sekretorni polovi usmjereni su prema ekvatoru stanice koja se dijeli, što osigurava aktivno sudjelovanje ove organele u formiranju srednje ploče i primarne stanične membrane.
Asimetrični polaritet karakterističan je za specijalizirane stanice. Dakle, u zrelom jajetu, jezgra je pomaknuta prema polu usmjerenom prema halazi ovula, a velika vakuola nalazi se na polu mikropilara.
Asimetrični polaritet je još izraženiji kod oplođene jajne stanice – zigote. U njemu se jezgra, okružena tubulima endoplazmatskog retikuluma, nalazi na halazalnoj strani stanice. Ovdje su gušće raspoređeni elementi citoskeleta, povećava se optička gustoća citoplazme, a povećan je sadržaj enzima, fitohormona i drugih fiziološki aktivnih tvari. Njegov funkcionalno aktivniji dio postaje apikalni, a suprotni dio postaje bazalni.
Iz vršne stanice tijekom razvoja sjemenog zametka nastaje embrionalna stabljika s konusom rasta i primordijalnim listićima. Bazalna stanica tvori visak, a kasnije spinalni korijen.
Zbog polariteta stanica embrija u razvoju, zrelo sjeme također postaje polarizirano. Na primjer, u sjemenu pšenice s jedne strane nalazi se endosperm, a s druge embrij, u kojem su vršni meristem stabljike i vrh korijena na različitim polovima.
Za razliku od stanica, polaritet biljnih organa je univerzalniji. Manifestira se u strukturi izdanka i korijenskog sustava, kao i njihovih komponenti. Primjerice, bazalni i vršni dio mladice razlikuju se morfološki, anatomski, histološki, biokemijski i funkcionalno. Kod mnogih cvjetnica na vrhu izboja nalazi se vršni pupoljak u kojem se nalazi najvažnije morfogenetsko središte biljke – konus rasta. Zahvaljujući njegovoj aktivnosti formiraju se primordije lišća, bočni aksilarni pupoljci, čvorovi i internodije. Diferencijacija stanica primarnog meristema dovodi do pojave primarnih integumentarnih tkiva, histoloških elemenata primarnog korteksa i središnjeg cilindra.
Vrh izdanka snažno je središte privlačenja. Ovdje se šalje glavni tok vode i mineralnih hranjivih elemenata otopljenih u njoj i organskih tvari sintetiziranih u korijenu. Preferencijalna opskrba vršnog pupoljka citokininom dovodi do apikalne dominacije. Stanice vrha apikalnog pupoljka aktivno se dijele, osiguravajući rast stabljike u duljinu i stvaranje novih listova i aksilarnih pupova na glavnoj osi. U ovom slučaju uočava se korelativna inhibicija razvoja aksilarnih pupoljaka.
Dobar primjer Morfološki polaritet mladice je građa stabljike pšenice. Kako se krećete odozdo prema gore, od bazalnog dijela izbojka do apikalnog dijela, internodije postaju duže, njihova debljina u srednjem dijelu prvo se povećava, a zatim se postupno smanjuje. U lišću gornjeg sloja, omjer širine lisne ploče i njegove duljine znatno je veći nego u lišću donjeg sloja. Mijenja se i položaj lišća u prostoru. U pšenici se listovi donjeg sloja spuštaju, listovi srednjeg sloja nalaze se gotovo plagiotropno, tj. paralelno s površinom zemlje, a zastava (gornji) listovi teže ortotropnom - gotovo okomitom - položaju.
Polaritet izdanka jasno je vidljiv na anatomskoj razini. Kod vrsta pšenice subklasni internodij, u usporedbi s nižim internodijama, karakterizira manji promjer, manje vaskularnih snopova i bolje razvijen asimilacijski parenhim.
U dikotilnim biljkama anatomska polarnost izdanka je pojačana zbog pojave sekundarnih bočnih obrazovnih tkiva - kambija i felogena. Felogen daje felem sekundarnog pokrovnog tkiva, koji se, zasićen suberinom, pretvara u čep. Kod vrsta drveća aktivnost felogena dovodi do stvaranja tercijarnog pokrovnog kompleksa – kore. Kambij osigurava prijelaz u sekundarnu anatomsku strukturu stabljike ispod razine vršnog pupoljka.
Korijenje, kao i druge biljne organe, također karakterizira strukturna i funkcionalna polarnost. Zbog pozitivnog geotropizma, bazalni dio korijena nalazi se na površini tla. Izravno je povezan s vratom korijena - mjestom gdje korijen prelazi u stabljiku. Apikalni dio korijena obično je zakopan u tlo. Prolazi kroz međusobno povezane procese rasta i razvoja. Dioba stanica vrha meristema osigurava linearni rast korijena. A kao rezultat diferencijacije stanica nastaju kvalitativno nove strukture. Na stanicama epibleme pojavljuju se korijenove dlačice. Od stanica peribleme nastaju elementi primarne kore, a od plerome pericikl i provodni elementi središnjeg cilindra. Ovakav dizajn mladih dijelova korijena osigurava aktivnu apsorpciju vode i minerala, kao i njihovu opskrbu višim dijelovima korijena. Funkcije bazalnog dijela korijena su nešto drugačije. Dakle, kod višegodišnjih dikotiledona bazalni dio korijena obavlja transportne, potporne i skladišne funkcije. Anatomska struktura odgovara izvedbi ovih funkcija. Ovdje su vodljiva tkiva bolje razvijena, kora se formira djelovanjem kambija, a pokrovno tkivo predstavlja pluto. Polarnost strukture korijena osigurava raznolikost njegovih funkcija.
Polaritet je također karakterističan za reproduktivne organe biljaka. Dakle, cvijet, kao modificirani skraćeni izdanak, zadržava znakove polariteta izdanka. Na njemu su u prirodnom nizu smješteni dijelovi cvijeta: čaška, vjenčić, androcej i ginecej. Ovakav raspored doprinosi boljem hvatanju peluda tučkom tučka, a također osigurava zaštitu generativnih dijelova cvijeta vegetativnim dijelovima. Vrlo je indikativan polaritet cvatova. Kod neodređenih cvatova najprije se formiraju cvjetovi bazalnog dijela. Oni dosežu velike veličine te tvore plodove s dobro razvijenim sjemenkama. Konkretno, krupnije sjemenke suncokreta nastaju u perifernom dijelu košare, au složenom klasu pšenice najbolje sjemenke nastaju u prvim cvjetovima klasića. Kasnije se formiraju cvjetovi vršnog dijela cvatova. Plodovi i sjeme dobiveni od njih su sitniji, a sjetvena kvaliteta slabija.
Dakle, polaritet je važna strukturna, funkcionalna i biokemijska značajka biljaka, koja ima adaptivno značenje i koja se mora uzeti u obzir u agronomskoj praksi.
1.2. Simetrija
Svijet oko nas karakterizira cjelovitost i skladan red. U obližnjem svemiru, položaj, masa, oblik i putanja objekata u Sunčevom sustavu su skladni. Na Zemlji su harmonične sezonske i dnevne promjene najvažnijih fizikalnih parametara uvjeta postojanja živih organizama, koje karakterizira i skladna usklađenost strukture i funkcija. Mudraci Pitagorine škole vjerovali su da je harmonija “način usklađivanja mnogih dijelova, uz pomoć kojih se oni ujedinjuju u cjelinu”. Simetrija je odraz harmonije u prirodi. Prema definiciji Yu.A. Urmantsevska simetrija je kategorija koja označava očuvanje značajki objekata u odnosu na njihove promjene. Utilitaristički rečeno, simetrija predstavlja ujednačenost strukture i međusobnog rasporeda sličnih sastavnih dijelova jedinstvene cjeline. Simetrija je svojstvena i mineralima i živim bićima. Međutim, oblici simetrije i stupanj njihove manifestacije u različitim objektima značajno variraju.
Bitna značajka simetrije objekata različitog podrijetla je njezina korelativna priroda. Usporedba struktura provodi se na nekoliko karakterističnih točaka. Točke figure koje daju istu sliku gledajući lik s različitih strana nazivamo jednakima. To mogu biti ekvidistantne točke na pravoj liniji, točke sjecišta stranica jednakokračnog trokuta i kvadrata, plohe poliedra, točke na kružnici ili na plohi lopte itd.
Ako na slici za neku točku X ne postoji druga jednaka točka, tada se ona naziva singularnom. Figure koje imaju jednu posebnu točku i više jednakih točaka nazivaju se rozete. Oblici koji ne sadrže jednake točke smatraju se asimetričnima.
Kroz singularne točke prolaze osi i ravnine simetrije. U jednakokračnom trokutu mogu biti tri, u kvadratu - četiri, u jednakostraničnom peterokutu - pet ravnina simetrije. Prema tome, dvije zrake simetrije formiraju se na ravnoj liniji, trokut karakterizira trozračna simetrija, kvadrat je karakteriziran četverozračnom simetrijom, a krug je karakteriziran višezračnom simetrijom.
Kretanja lika, uslijed kojih se svaka točka zamjenjuje jednakom točkom, a svaka singularna točka ostaje na mjestu, nazivaju se transformacijama simetrije, a figure za koje je transformacija simetrije dopuštena smatraju se simetričnima.
Najčešći oblici transformacije simetrije su:
1. Refleksija - kretanje svake točke koja se nalazi na određenoj udaljenosti od fiksne ravnine duž ravne crte okomite na ovu ravninu do iste udaljenosti s druge strane (na primjer, zrcalna simetrija zigomorfnog cvijeta graška);
2. Rotacija – pomicanje svih točaka pod određenim kutom oko fiksne osi (npr. višezračna simetrija aktinomorfnog cvijeta trešnje);
3. Paralelni prijenos npr. položaja metamera na mladici.
Oblici transformacije simetrije nisu identični i ne mogu se reducirati jedan na drugi. Dakle, u slučaju refleksije, ravnina ostaje nepomična; pri okretanju – ravno (os); a kod paralelnog prijenosa niti jedna točka ne ostaje na mjestu.
Simetrični likovi s više jednakih točaka mogu imati različit broj singularnih točaka. Dakle, utičnica ima jednu posebnu točku. Glavni oblik transformacije simetrije za njega je rotacija. Za figure koje nemaju posebnih točaka karakteristična je paralelna translacija, odnosno pomak. Takve figure se konvencionalno nazivaju beskonačnim.
2.2.1. Značajke manifestacije simetrije u biljkama. Simetrija biljaka razlikuje se od simetrije kristala u nizu značajki.
1) Simetrija biljaka određena je ne samo simetrijom molekula koje tvore njihove stanice, već i simetrijom uvjeta okoliš, u kojem se odvija razvoj biljaka.
Posebnim slučajevima simetrije staništa može se smatrati ujednačenost sastava korijenskog sloja tla, ujednačena raspodjela vode i elemenata mineralne prehrane u tlu i jednaka udaljenost biljaka koje rastu u blizini. Ove okolnosti moraju se uzeti u obzir pri uzgoju kultiviranih biljaka.
2) Biljke, kao i drugi živi organizmi, nemaju apsolutnu istovjetnost elemenata svojih sastavnih dijelova. To je prvenstveno određeno činjenicom da uvjeti za nastanak ovih dijelova nisu apsolutno identični. S jedne strane, to je zbog različitih vremena u kojima stožac rasta izolira različite metamere, uključujući primordije listova, pupova, čvorova i internodija. Isto vrijedi i za generativne organe. Tako su rubni cjevasti cvjetovi u suncokretovoj košari uvijek veći od cvjetova u središnjem dijelu cvata. S druge strane, tijekom vegetacije uočavaju se redovite i slučajne promjene uvjeta razvoja. Na primjer, temperatura zraka i tla, osvjetljenje i spektralni sastav svjetlosti statistički se mijenjaju tijekom vegetacijske sezone. Lokalne promjene u opskrbi hranjivim tvarima u tlu i učinci bolesti i štetnika mogu biti nasumični.
Odsutnost apsolutne istovjetnosti sastavnih dijelova od velike je važnosti u životu biljaka. Pojavljujuća heterogenost jedan je od mehanizama pouzdanosti ontogeneze.
3) Biljke karakterizira supstancijalna, prostorna i vremenska simetrija.
A. Supstancijalna simetrija sastoji se u točnom ponavljanju oblika i linearnih parametara konstrukcija, kao iu točnoj promjeni tih parametara. Karakterističan je za kristale anorganske tvari, molekule organskih spojeva, stanične organele, anatomske strukture i organi biljaka. Visoka razina Molekule DNA razlikuju se po simetriji tvari. Kada je vlažnost pripravaka blizu fiziološke, molekula DNA je u B-formi i karakterizirana je jasnim ponavljanjem parametara svojih sastavnih elemenata. Svaki zavoj molekule DNA sadrži 10 nukleotida; projekcija zavoja na os molekule je 34,6 Å (1 Å = 1·10–10 m); udaljenost projekcije između susjednih nukleotida je 3,4 Å, a linearna udaljenost je 7 Å; promjer molekule kada je orijentirana duž atoma fosfora je blizu 20 Å; Promjer većeg žlijeba je približno 17 Å, a sporednog žlijeba 11 Å.
Supstancijalnu simetriju biljaka karakterizira ne samo točno ponavljanje strukturnih parametara, već i njihova pravilna promjena. Na primjer, kod smreke je smanjenje promjera debla po 1 m duljine pri kretanju od bazalnog dijela prema vrhu relativno konstantno. Ljubičasta kana (Canna violacea) ima cvijet koji se tradicionalno smatra asimetričnim. Njegove čašice, latice i staminodije su različite veličine. Međutim, promjena linearnih dimenzija ovih cvjetnih članova ostaje relativno konstantna, što je znak simetrije. Samo se ovdje umjesto zrcalne simetrije razvijaju drugi oblici simetrije.
B. Prostorna simetrija sastoji se od prirodnog ponavljanja prostornog rasporeda sličnih komponenti u biljkama. Prostorna simetrija je raširena u biljnom svijetu. Karakterističan je za raspored pupova na izdanku, cvjetova u cvatu, cvjetnih članova na prihvatu, peluda u prašniku, ljuski u češerima golosjemenjača i mnoge druge slučajeve. Poseban slučaj prostorne simetrije je uzdužna, radijalna i mješovita simetrija stabljike.
Uzdužna simetrija se javlja tijekom paralelnog prijenosa, tj. prostorno ponavljanje metamera u strukturi izdanka. Određuje se strukturnom sličnošću sastavnih dijelova izdanka - metamera, kao i sukladnošću duljine internodija s pravilom "zlatnog presjeka".
Složeniji slučaj prostorne simetrije je raspored listova na mladici, koji može biti kovrdžast, nasuprotan ili naizmjeničan (spiralan).
Radijalna simetrija nastaje kada se os strukture kombinira s ravninama simetrije koje prolaze kroz nju. Radijalna simetrija je široko rasprostranjena u prirodi. Osobito je karakterističan za mnoge vrste dijatomeja, presjek stabljike viših sporišnih biljaka, golosjemenjača i kritosjemenjača.
Radijalna simetrija izdanka je višeelementna. Karakterizira položaj sastavnih dijelova i može se izraziti različitim dimenzionalnim pokazateljima: udaljenošću vaskularnih snopova i drugih anatomskih struktura od posebne točke kroz koju prolazi jedna ili više ravnina simetrije, ritmičkom izmjenom anatomskih struktura, kutom divergencije. , koji pokazuje pomak osi jedne strukture u odnosu na različitu os. Na primjer, u cvijetu trešnje pet vjenčića je prostorno simetrično raspoređeno. Mjesto pričvršćivanja svakog od njih na posudu nalazi se na istoj udaljenosti od središta cvijeta, a kut divergencije bit će jednak 72º (360º: 5 = 72º). U cvijetu tulipana, kut divergencije svake od šest latica je 60º (360º: 6 = 60º).
C. Vremenska simetrija biljaka izražava se u ritmičkom ponavljanju u vremenu procesa morfogeneze i drugih fizioloških funkcija. Na primjer, stožac rasta primordija lišća izoliran je u više ili manje jednakim vremenskim intervalima, koji se nazivaju plastokroni. Sezonske promjene u vitalnim procesima u višegodišnjim polikarpskim biljkama ponavljaju se vrlo ritmički. Vremenska simetrija odražava prilagodbu biljaka dnevnim i sezonskim promjenama okolišnih uvjeta.
4) Simetrija biljaka dinamički se razvija tijekom ontogeneze i dostiže svoj maksimalni izražaj tijekom spolnog razmnožavanja. Kod biljaka je indikativan primjer u tom smislu stvaranje radijalne simetrije izdanka. U početku su stanice konusa rasta više ili manje homogene i nediferencirane. Stoga je morfološki teško identificirati stanicu ili skupinu stanica kroz koju bi se mogla povući ravnina simetrije.
Kasnije stanice tunike tvore protodermis iz kojeg nastaje epidermis. Iz glavnog meristema razvijaju se asimilirajuća i skladišna tkiva, primarna mehanička tkiva i jezgra. U perifernoj zoni stvaraju se niti uskih i dugih prokambijskih stanica iz kojih će se razviti provodna tkiva. Kada je prokambij potpuno položen, iz njega nastaju kontinuirani slojevi floema i ksilema. Ako je prokambij položen u obliku užeta, tada se iz njega formiraju zasebni provodni snopovi. Kod drvenastih biljaka, višegodišnja, sezonski promjenjiva funkcionalna aktivnost kambija dovest će do stvaranja sekundarne kore i godišnjih drvenih godova, što će poboljšati radijalnu simetriju stabljike.
5) Simetrija biljaka se mijenja tijekom evolucije. Evolucijske transformacije simetrije imaju velika vrijednost u razvoju organskog svijeta. Dakle, pojava bilateralne simetrije bila je velika morfofiziološka prilagodba (aromorfoza), koja je značajno povećala razinu organizacije životinja. U biljnom carstvu promjena broja osnovnih tipova simetrije i njihovih izvedenica povezana je s pojavom višestaničnosti i pojavom biljaka na kopnu. Kod jednostaničnih algi, posebice dijatomeja, radijalna je simetrija široko rasprostranjena. U višestaničnim organizmima pojavljuju se raznih oblika paralelni prijenos, kao i mješoviti oblici simetrije. Na primjer, u karofitnim algama, radijalna simetrija poprečnog presjeka talusa kombinirana je s prisutnošću osi prijenosa metamera.
Evolucija provodnog sustava bila je od velike važnosti u formiranju simetrije viših biljaka. Radijalno simetrična protostela stabljike primitivnih oblika zamijenjena je kompleksom bilateralno simetričnih (monosimetričnih) kolateralnih snopova koji tvore eustelu i osiguravaju radijalnu simetriju stabljike u angiospermi.
Opći trend u evoluciji simetrije u živim organizmima, uključujući i biljke, je smanjenje razine simetrije. To je zbog smanjenja broja osnovnih tipova simetrije i njihovih izvedenica. Tako evolucijski ranije cvjetnice (familija Magnoliaceae, familija Ranunculaceae i dr.) karakteriziraju polinomijalni, slobodni, spiralno raspoređeni cvjetni dijelovi. Ispada da je cvijet aktinomorfan, tj. polisimetričan. Formule takvog cvijeća obično su: Å Ca ¥ Co ¥ A ¥ G ¥ . Tijekom evolucije uočava se smanjenje broja članova cvijeta, kao i njihovo spajanje, što uvijek dovodi do smanjenja broja ravnina simetrije.
U evolucijski mladim obiteljima, na primjer, u Lamiaceae ili Poagrass, cvjetovi postaju zigomorfni (monosimetrični). Disimetrizacija cvjetova očito ima adaptivno značenje povezano s poboljšanjem metoda oprašivanja. Ovo je često bilo olakšano konjugiranom evolucijom cvijeta i oprašivača - insekata i ptica.
Specijalizacija simetrije cvjetova u cvatu stekla je veliku važnost. Tako u košari suncokreta sterilni rubni zigomorfni cvjetovi imaju veliki lažni jezik žuta boja, koju čine tri srasle latice. Fertilni cvjetovi u središnjem dijelu cvata su aktinomorfni, tvore ih pet malih jednakih latica sraslih u cijev. Drugi primjer su predstavnici obitelji celera. U njihovom složenom cvjetištu, rubni cvjetovi su slabo zigomorfni, dok ostali cvjetovi ostaju tipično aktinomorfni.
Evolucija strukture i simetrije biljaka nije jednostavna. Pretežnu ulogu disimetrizacije u određenim stupnjevima evolucije zamjenjuje i nadopunjuje simetrizacija.
Dakle, simetrija biljaka i njihovih komponenti vrlo je višestruka. Povezan je sa simetrijom njegovih sastavnih elemenata na molekularnoj, staničnoj, histološkoj, anatomskoj i morfološkoj razini. Simetrija se dinamički razvija tijekom ontogeneze i filogeneze i osigurava povezanost biljaka s okolišem.
1.3. Metamerija
Važna morfofiziološka prilagodba biljaka je metamerija, što je prisutnost ponavljajućih elementarnih konačnih struktura ili metamera u sustavu cijelog organizma. Metamerna struktura osigurava višestruko ponavljanje sastavnih dijelova izdanka i stoga je jedan od mehanizama pouzdanosti ontogeneze. Metamerna struktura karakteristična je za različite sistematske skupine biljaka. Poznat je iz harofitnih algi, preslica i drugih viših sporišnih biljaka, golosjemenjača, kopnenih i vodenih kritosjemenjača. Funkcionalno različiti dijelovi biljaka su metameri – vegetativni i generativni. Uz sudjelovanje metamerno raspoređenih organa, u biljkama se formira metamerni sustav. Poseban slučaj nastanka metamernih sustava je grananje.
Metamera vegetativne zone izdanka angiosperma uključuje list, čvor, internodij i bočni aksilarni pupoljak, koji se nalazi u podnožju internodija na strani suprotnoj od mjesta pričvršćivanja lišća. Ovaj pupoljak je prekriven listom prethodnog metamera. Metameri generativne zone vrlo su raznoliki. Na primjer, kod pšenice, metameri složenog klasa sastoje se od segmenta stabljike klasa i na njega pričvršćenog klasića. Ponekad biljke imaju prijelaznu zonu izdanaka. Kod nekih vrsta i sorti pšenice može se prikazati ljuskama nerazvijenih klasića.
Metamerija biljaka je morfološki izraz specifičnosti njihova rasta i morfogeneze, koji se odvijaju ritmički, u obliku ponavljanja podređenih ciklusa lokaliziranih u žarištima meristema. Ritam stvaranja metamera neraskidivo je povezan s periodičnošću procesa rasta karakterističnih za biljke. Stvaranje metamera primarni je morfogenetski proces u razvoju biljaka. On čini osnovu za kompliciranje organizacije u ontogenezi i odražava proces polimerizacije, koji je jedan od mehanizama evolucije viših biljaka.
Formiranje i razvoj metamera osigurava se prvenstveno funkcijom apikalnog i interkalarnog meristema.
U vegetativnom pupoljku, kao iu embriju klijajućeg sjemena, kao rezultat mitotske diobe, povećava se volumen konusa rasta embrionalne peteljke. Naknadna aktivna dioba stanica u perifernoj zoni konusa rasta dovodi do stvaranja lisnog primordija - lisnog primordija i insercijskog diska. U to vrijeme meristematska aktivnost tunike i središnje meristematske zone nešto opada, ali se stanice insercijskog diska aktivno dijele. Gornji dio diska je mjesto pričvršćivanja primordija lista i, kako raste u debljini, iz njega se formira čvor. Internodij se razvija iz donjeg dijela insercijskog diska. Ovdje se formira rudimentarna kvržica bočnog aksilarnog pupoljka na strani nasuprot središnjoj žilici primordijalnog lista. Zajedno, primordij lista, insercijski disk i primordij pupoljka čine rudimentarni metamer.
Kako je formiranje embrionalnog metamera završeno, aktivnost stanične diobe u tunici i središnjoj meristematskoj zoni ponovno raste. Volumen glatkog dijela stošca rasta ponovno se povećava, dosežući maksimum prije pokretanja sljedećeg lisnog primordija. Tako se na apikalnom dijelu prethodne rudimentarne metamere počinje formirati nova. Ovaj proces je genetski određen i ritmički se ponavlja mnogo puta. U tom će slučaju prvi formirani metamer biti smješten u bazalnom dijelu izdanka, a ontogenetski najmlađi bit će smješten u vršnom dijelu. Nakupljanje broja metamernih primordija u bubregu T.I. Serebrjakova je to nazvala sazrijevanjem. Za označavanje maksimalnog broja metamera taloženih u bubregu, predložila je izraz "kapacitet bubrega".
Rast i razvoj internodija, kao sastavnica metamera, uvelike je određen aktivnošću interkalarnog meristema. Dioba stanica ovog meristema i izduživanje njihovih derivata dovodi do izduživanja internodija.
Metameri imaju broj karakteristične značajke, koji im omogućuju da osiguraju strukturni i funkcionalni integritet biljnog organizma.
1) Polaritet metamera. Svaki metamer ima bazalni i apikalni dio. Izraz "apikalni dio" sugerira da gornji dio Metamer ili sadrži apikalni meristem ili je orijentiran prema vrhu.
Bazalni i apikalni dio razlikuju se po morfološkim, histološko-anatomskim i fiziološko-biokemijskim karakteristikama. Na primjer, kod pšenice, kada se kreće od bazalnog prema vršnom dijelu internodija, debljina stabljike, promjer medularne lakune i debljina perifernog sklerenhimskog prstena prvo se povećavaju, a zatim postupno smanjuju; značajno se povećava broj stanica u nitima asimilacijskog parenhima; Radijalni promjer vaskularnih snopova se smanjuje, kao i broj žila u ksilemu.
2) Simetrija metamera. Simetrija biljnih organa osigurava se simetrijom metamera koji ih tvore, a koja nastaje kao rezultat specifičnog slijeda stanične diobe u stošcu rasta.
3) Heterokroničnost nastanka metamera. Metameri su izolirani u različito vrijeme, naizmjenično, konusom rasta. Stoga su prve metamere bazalnog dijela mladice ontogenetski starije, a posljednje metamere vršnog dijela mlađe. Morfološki se razlikuju po parametrima lista, duljini i debljini internodija. Na primjer, kod žitarica, listovi gornjeg sloja su širi od listova donjeg sloja, a podšiljci su duži i tanji od donjih.
4) Varijabilnost karakteristika metamera. Ontogenetski mlađe metamere imaju manju amplitudu varijabilnosti svojstva. Stoga se anatomske karakteristike internodija podšiljka mogu koristiti s većom točnošću za identifikaciju sorti usjevi žitarica te izrada uzgojnih programa.
5) Optimalni dizajn metamera. Optimalna biološka struktura je ona koja zahtijeva minimalnu količinu organske tvari za izgradnju i održavanje. Budući da je građa biljaka neraskidivo povezana s njihovom funkcijom, kriterij optimalnosti može biti prilagodljivost biljaka uvjetima uzgoja i omjer njihove sjemenske produktivnosti i mase vegetativnih organa. Optimalan dizajn cijelog postrojenja osiguran je optimalnim dizajnom njegovih sastavnih metamera.
6) Kompletna biljka je polimerni sustav sastavljen od skupa metamera. Polimernost se očituje na svim razinama organizacije biljnog organizma. Dakle, u strukturi DNA postoji mnogo ponavljanja gena; Kariotip mnogih biljaka predstavljen je diploidnim ili poliploidnim skupom kromosoma; plastidi, mitohondriji, ribosomi i drugi organeli nalaze se u velikim količinama u stanici. Odrasla biljka je podređeni skup izdanaka, korijena i rasplodnih organa koji imaju metamernu strukturu. Metamerni sustav osigurava visoku produktivnost biljaka i značajno povećava pouzdanost njihove ontogeneze.
27. 4.1. Primarna anatomska građa korijena
Značajke primarne strukture jasno se očituju u uzdužnim i poprečnim presjecima vrha korijena.
Na uzdužnom presjeku vrha korijena mogu se razlikovati četiri zone
Zona korijenske kapice pokriva apikalne meristeme korijena. Sastoji se od živih stanica. Njihov površinski sloj stalno se ljušti i oblaže prolaz po kojem se kreće korijen. Stanice ljuštenja također proizvode sluz, što olakšava kretanje vrha korijena u tlu. Stanice središnjeg dijela klobuka ili kolumele sadrže škrobna zrnca koja pridonose geotropnom rastu korijena. Stanice kapice stalno se obnavljaju zbog diobe stanica posebnog obrazovnog tkiva - kaliptrogena, karakterističnog za jednosupnice.
I – zona korijenske kapice; II – zona rasta; III – zona korijenskih dlačica; IV – prostor za držanje. 1 – epiblema, 2 – pericikl, 3 – endoderm, 4 – primarni korteks, 5 – egzodermis, 6 – središnji cilindar, 7 – korijenska dlaka; 8 – formiranje bočnog korijena.
Zona rasta sastoji se od dvije podzone. U podzoni diobe dolazi do rasta korijena zbog aktivne mitotske diobe stanica. Na primjer, u pšenici, udio stanica koje se dijele (mitotski indeks) je 100-200 ppm. Podzona diobe je vrijedan materijal za citogenetička istraživanja. Ovdje je prikladno proučavati broj, makro- i mikrostrukturu kromosoma. U podzoni elongacije smanjuje se meristematska aktivnost stanica, ali zbog određene ravnoteže fitohormona, prvenstveno auksina i citokinina, dolazi do rasta korijena zbog aksijalne elongacije mladih stanica.
Zona apsorpcije se s pravom može nazvati zonom korijenske dlake, kao i zonom diferencijacije, budući da se tu formiraju epiblema, primarni korteks i središnji cilindar.
Epiblema je posebno, stalno obnavljano pokrovno tkivo koje se sastoji od dvije vrste stanica. Iz trihoblasta razvijaju se tankostjedne korijenske dlake duljine 1-3 mm, zahvaljujući kojima se apsorbira voda i u njoj otopljene tvari. Korijenove dlake su kratkotrajne. Žive 2-3 tjedna, a zatim se deskvamiraju. Atrihoblasti ne stvaraju korijenske dlake i obavljaju pokrovnu funkciju.
Primarna kora regulira protok vode u provodna tkiva središnjeg cilindra. Kada se formira središnji cilindar, prvo nastaju tkiva protofloema, kroz koja u vrh korijena ulaze organske tvari potrebne za njegov rast. Više od korijena, u razini prvih korijenovih dlačica, pojavljuju se vaskularni elementi protoksilema. Početna vaskularna tkiva razvijaju se u primarni floem i primarni ksilem radijalnog vaskularnog snopa.
Provodna zona zauzima najveći dio korijena. Obavlja brojne funkcije: transport vode i u njoj otopljenih mineralnih i organskih tvari, sintezu organskih spojeva, skladištenje hranjivih tvari itd. Provodna zona završava vratom korijena, t.j. prijelazna točka između korijena i stabljike. Kod dvosupnih angiosjemenjača na početku ove zone nalazi se prijelaz iz primarne u sekundarnu anatomsku građu korijena.
Presjek u području korijenovih dlačica otkriva strukturne i topografske značajke primarne anatomske građe korijena (slika 2). Na površini korijena nalazi se epiblema s korijenovim dlačicama, uz pomoć kojih se upija voda i u njoj otopljene tvari. Za razliku od epidermisa, epiblema nije prekrivena kutikulom i nema stomata. Ispod njega je primarni korteks, koji se sastoji od egzoderma, mezoderma i endoderma.
1 – zone vrhova korijena; 2 – 6 – poprečni presjeci na različitim razinama; Vx - sekundarni ksilem; Vf – sekundarni floem; K – kambij; Mx – metaksilem; Mf – metafloem; Pd – protoderm; Px – protoksilem; Prd – periderm; Pf – protofloem; Pc – pericikl; Rd – rizoderm; Ex – egzodermis; En – endoderm; C – poklopac.
Egzodermalne stanice su gusto naborane i imaju zadebljale membrane. Egzoderm osigurava apoplastičnu opskrbu vodom dubljih slojeva korteksa, a također daje snagu korijenu s površine.
Srednji sloj korteksa, mezoderm, sastoji se od stanica parenhima tankih stijenki i ima labavu strukturu zbog prisutnosti brojnih međustaničnih prostora kroz koje se voda kreće do egzoderma.
Endoderm, ili unutarnji sloj primarne kore, sastoji se od jednog reda stanica. Sastoji se od kaveza s kasparskim pojasevima i prolaznih kaveza. Stanice s kasparskim pojasom imaju zadebljane lateralne (radijalne) i tangencijalne (krajnje) membrane okrenute prema središnjem cilindru. Ta celulozna zadebljanja impregnirana su ligninom i stoga ne mogu propuštati vodu. Nasuprot tome, prolazne stanice su tankih stijenki i smještene nasuprot zrakama ksilema. Kroz prolazne ćelije voda ulazi u radijalni vodljivi snop središnjeg cilindra.
Središnji cilindar korijena ili stele sastoji se od nekoliko slojeva stanica. Neposredno ispod endodermisa nalazi se jedan ili više redova pericikličnih stanica. Od njih, tijekom prijelaza na sekundarnu strukturu korijena u dikotilnim angiospermama i gimnospermama, formiraju se interfascikularni kambij i felogen (plutasti kambij). Osim toga, periciklične stanice sudjeluju u formiranju bočnih korijena. Iza pericikla nalaze se stanice radijalnog vaskularnog snopa.
Vaskularni snop nastaje iz prokambija. Najprije nastaju stanice protofloema, a zatim u razini prvih korijenovih dlačica - stanice protoksilema. Stanice provodnih tkiva nastaju egzarhijski, tj. od površine grede, a potom se razvijaju u centripetalnom smjeru. U ovom slučaju, najranije, veće posude nalaze se u središtu ksilema, a mlađe, manjeg promjera, nalaze se na periferiji ksilemske zrake. Stanice floema nalaze se između ksilemskih zraka.
Broj ksilemskih zraka ovisi o sistematskom položaju biljaka. Na primjer, neke paprati mogu imati samo jednu zraku ksilema i jedan dio floema. Tada se punđa zove monarch. Mnoge dikotiledone angiosperme karakteriziraju diarhični snopovi s dvije ksilemske zrake. Osim toga, imaju biljke s tri-, tetra- i pentarhijskim snopovima. Kod monokotiledonih angiospermi tipični su poliarhalni vaskularni snopovi s višezračenim ksilemom.
Struktura radijalnog vaskularnog snopa utječe na način pokretanja lateralnih korijena. U diarhnom korijenu nastaju između floema i ksilema, u triarhnom i tetrarhnom korijenu nasuprot ksilemu, a u poliarhnom korijenu nasuprot floemu.
Stanična struktura epibleme i primarnog korteksa osigurava pojavu korijenskog pritiska vode. Vodu apsorbiraju korijenove dlake, ostavlja ih u staničnoj membrani egzoderma, zatim ulazi u međustanične prostore mezoderma, a iz njih kroz prolazne stanice tankih stijenki u žile radijalnog vaskularnog snopa. Budući da korijenske dlačice imaju veću ukupnu površinu od prolaznih stanica, brzina kretanja vode, a time i njezin pritisak, povećava se kako se približava žilama. Razlika u tlaku koja nastaje predstavlja pritisak korijena, što je jedan od mehanizama protoka vode u stabljiku i druge organe biljke. Drugi važan mehanizam kretanja vode je transpiracija.
28. 4.2. Sekundarna anatomska građa korijena
Kod dvosupnih angiosjemenjača i golosjemenjača primarna anatomska struktura korijena u zoni provođenja nadopunjena je strukturama sekundarnog podrijetla, koje nastaju zbog nastanka i meristematske aktivnosti sekundarnih bočnih obrazovnih tkiva - kambija i felogena (pluta kambija) (Sl. 3). Prilikom prelaska na sekundarnu strukturu, sljedeće značajne promjene događaju se temeljno.
A. Pojava u središnjem cilindru korijena kambija i sekundarnog ksilema i sekundarnog floema stvorenog njime, koji su u smislu kapaciteta mnogo bolji od elemenata izvornog radijalnog vaskularnog snopa.
Ovaj tip pripada složenim tkivima i sastoji se od različito diferenciranih stanica. Osim samih vodljivih elemenata, tkivo sadrži mehaničke, ekskretorne i skladišne elemente. Provodna tkiva ujedinjuju sve biljne organe u jedinstveni sustav. Postoje dvije vrste provodnih tkiva: ksilem i floem (grč. xylon - stablo; phloios - kora, ličje). Imaju i strukturne i funkcionalne razlike.
Provodne elemente ksilema čine mrtve stanice. Oni vrše prijenos vode i tvari otopljenih u njoj na velike udaljenosti od korijena do lišća. Provodni elementi floema čuvaju živi protoplast. Oni obavljaju transport na velike udaljenosti od fotosintetskog lišća do korijena.
Obično se ksilem i floem nalaze u tijelu biljke određenim redoslijedom, tvoreći slojeve ili vaskularne snopove. Ovisno o građi, postoji nekoliko vrsta vaskularnih snopova, koji su karakteristični za pojedine skupine biljaka. Kolateralni otvoreni snop između ksilema i floema sadrži kambij, koji osigurava sekundarni rast. U bikolateralnom otvorenom snopu, floem se nalazi u odnosu na ksilem s obje strane. Zatvoreni snopovi ne sadrže kambij, pa stoga nisu sposobni za sekundarno zadebljanje. Možete pronaći još dvije vrste koncentričnih snopova, gdje ili floem okružuje ksilem, ili ksilem okružuje floem.
Xylem (drvo). Razvoj ksilema u višim biljkama povezan je s osiguravanjem izmjene vode. Budući da se voda neprestano uklanja kroz epidermu, istu količinu vlage biljka mora apsorbirati i dodati organima koji provode transpiraciju. Treba uzeti u obzir da bi prisutnost živog protoplasta u stanicama koje provode vodu uvelike usporila transport; mrtve stanice ovdje su funkcionalnije. Međutim, mrtva stanica nema čvrstoću, pa stoga mehanička svojstva mora imati školjku. Napomena: turgescencija - stanja biljne stanice, tkiva i organa, za koje? postaju elastične zbog pritiska staničnog sadržaja na njihove elastične membrane. Doista, provodni elementi ksilema sastoje se od mrtvih stanica izduženih duž osi organa s debelim lignificiranim školjkama.
U početku se ksilem formira iz primarnog meristema - prokambija, koji se nalazi na vrhovima aksijalnih organa. Najprije se diferencira protoksilem, zatim metaksilem. Poznata su tri tipa stvaranja ksilema. U egzarhičnom tipu elementi protoksilema prvo se pojavljuju na periferiji prokambijevog snopa, zatim se elementi metaksilema pojavljuju u središtu. Ako proces ide u suprotnom smjeru (tj. od centra prema periferiji), onda je to endarhični tip. Kod mezarhijskog tipa ksilem se stvara u središtu prokambijskog snopa, nakon čega se taloži i prema središtu i prema periferiji.
Za korijen je karakterističan egzarhalni tip tvorbe ksilema, a za stabljike endarhični tip. Kod nisko organiziranih biljaka, metode formiranja ksilema vrlo su raznolike i mogu poslužiti kao sustavne karakteristike.
Neki? U biljkama (na primjer, jednosupnice), sve stanice prokambija diferenciraju se u provodna tkiva koja nisu sposobna za sekundarno zadebljanje. U drugim oblicima (na primjer, drvenastim), bočni meristemi (kambij) ostaju između ksilema i floema. Te se stanice mogu dijeliti, obnavljajući ksilem i floem. Taj se proces naziva sekundarni rast. U mnogima raste relativno stabilno klimatskim uvjetima, biljke, rast je stalan. U oblicima prilagođenim sezonskim klimatskim promjenama - periodično.
Glavne faze diferencijacije stanica prokambija. Njegove stanice imaju tanke membrane koje ih ne sprječavaju da se rastegnu tijekom rasta organa. Protoplast tada počinje odlagati sekundarnu ljusku. Ali ovaj proces ima različite značajke. Sekundarna ljuska nije taložena u kontinuiranom sloju, koji ne bi omogućio rastezanje stanice, već u obliku prstena ili spirale. Produljenje ćelije nije teško. U mladim stanicama, prstenovi ili zavoji spirale nalaze se blizu jedan drugoga. U zrelim stanicama, stanice se razilaze kao rezultat produljenja stanice. Prstenasta i spiralna zadebljanja ljuske ne ometaju rast, ali su mehanički inferiorna u odnosu na ljuske, gdje sekundarno zadebljanje tvori kontinuirani sloj. U tom smislu, nakon prestanka rasta, u ksilemu se formiraju elementi s kontinuiranom lignificiranom ljuskom (metaksilem). Treba napomenuti da sekundarno zadebljanje ovdje nije prstenasto ili spiralno, već točkasto, skalariformno, mrežasto.Njegove stanice nisu sposobne rastezati se i umiru unutar nekoliko sati. Taj se proces odvija na koordiniran način u obližnjim stanicama. U citoplazmi se pojavljuje veliki broj lizosoma. Zatim se raspadaju, a enzimi sadržani u njima uništavaju protoplast. Kada su poprečne stijenke uništene, stanice smještene u lancu jedna iznad druge tvore šuplju posudu. Većina angiospermi i neke? pteridofiti imaju krvne žile.
Provodna stanica koja se ne formira kroz perforacije u svojoj stijenci naziva se traheida. Kretanje vode kroz traheide odvija se manjom brzinom nego kroz posude. Činjenica je da kod traheida primarna ljuska nigdje nije prekinuta. Traheide međusobno komuniciraju kroz pore. Treba pojasniti da je kod biljaka pora samo udubljenje u sekundarnoj ljusci do primarne ljuske i nema prolaznih perforacija između traheida.
Najčešće su obrubljene pore. U njima kanal koji je okrenut prema staničnoj šupljini tvori proširenje – komoru pore. Najviše pore crnogorične biljke na primarnoj ljusci imaju zadebljanje - torus, koji je svojevrsni ventil i sposoban je regulirati intenzitet transporta vode. Pomicanjem torus blokira protok vode kroz poru, ali se nakon toga više ne može vratiti u prethodni položaj, vršeći jednokratnu radnju.
Pore su više ili manje okrugle, izdužene okomito na izduženu os (skupina tih pora nalikuje ljestvama; stoga se takva poroznost naziva stubištem). Kroz pore se transport odvija i u uzdužnom i u poprečnom smjeru. Pore su prisutne ne samo u traheidima, već iu pojedinačnim vaskularnim stanicama koje tvore krvnu žilu.
Sa stajališta evolucijske teorije, traheide predstavljaju prvu i glavnu strukturu koja provodi vodu u tijelu viših biljaka. Vjeruje se da su žile nastale iz traheida zbog razgradnje poprečnih stijenki između njih. Većina pteridofita i golosjemenjača nema žile. Njihovo kretanje vode odvija se kroz traheide.
U procesu evolucijskog razvoja plovila su nastala u različite grupe biljke opetovano, ali su najvažnije funkcionalno značenje dobile u kritosjemenjačama, u kojima? prisutni su uz traheide. Vjeruje se da im je posjedovanje naprednijeg transportnog mehanizma pomoglo ne samo da prežive, već i da postignu značajnu raznolikost oblika.
Ksilem je složeno tkivo, osim elemenata koji provode vodu sadrži i druge. Mehaničke funkcije izvode libriformna vlakna (latinski liber - ličje, forma - oblik). Prisutnost dodatnih mehaničkih struktura je važna jer su, unatoč zadebljanju, stijenke elemenata koji provode vodu još uvijek pretanke. Oni nisu u stanju sami podržati velike mase. višegodišnja biljka. Vlakna su se razvila iz traheida. Karakteriziraju ih manje veličine, lignificirane (lignificirane) školjke i uske šupljine. Na zidu se mogu naći pore bez granica. Ova vlakna ne mogu provoditi vodu, njihova glavna funkcija je podrška.
Ksilem također sadrži žive stanice. Njihova masa može doseći 25% ukupnog volumena drva. Budući da su te stanice okruglog oblika, nazivaju se drveni parenhim. U tijelu biljke parenhim je smješten na dva načina. U prvom slučaju, stanice su raspoređene u obliku okomitih niti - to je parenhim niti. U drugom slučaju, parenhim formira horizontalne zrake. Nazivaju se sržnim zrakama jer povezuju srž i korteks. Jezgra obavlja niz funkcija, uključujući skladištenje tvari.
Floem (ličje). Ovo je složeno tkivo jer ga čine različite vrste stanica. Glavne provodne ćelije nazivamo sitastim elementima. Provodne elemente ksilema čine mrtve stanice, dok u floemu zadržavaju živi, iako jako modificirani protoplast tijekom razdoblja funkcioniranja. Floem provodi odljev plastičnih tvari iz fotosintetskih organa. Sve žive biljne stanice imaju sposobnost provođenja organskih tvari. I stoga, ako se ksilem može naći samo u višim biljkama, tada se transport organskih tvari između stanica također provodi u nižim biljkama.
Ksilem i floem razvijaju se iz vršnih meristema. U prvoj fazi protofloem se formira u prokambijalnoj vrpci. Kako okolna tkiva rastu, rasteže se, a kada je rast završen, umjesto protofloema nastaje metafloem.
U različitim skupinama viših biljaka mogu se naći dvije vrste elementi sita. Kod pteridofita i golosjemenjača predstavljen je sitastim stanicama. Sitasta polja u ćelijama razbacana su duž bočnih stijenki. Protoplast zadržava donekle destruiranu jezgru.
Kod angiospermi se sitasti elementi nazivaju sitastim cijevima. Međusobno komuniciraju preko sitastih ploča. Zrele stanice nemaju jezgre. Međutim, uz sitastu cijev nalazi se stanica pratilica, nastala zajedno sa sitastom cijevi kao rezultat mitotske diobe zajedničke matične stanice (slika 38). Stanica pratilac ima gušću citoplazmu s velikim brojem aktivnih mitohondrija, kao i potpuno funkcionalnu jezgru, ogroman broj plazmodezmata (deset puta više od ostalih stanica). Prateće stanice utječu na funkcionalnu aktivnost sitastih stanica bez jezgre.
Struktura zrelih sitastih stanica ima neke osobitosti. Nema vakuole, pa je citoplazma jako ukapljena. Jezgra može biti odsutna (kod angiospermi) ili u naboranom, funkcionalno neaktivnom stanju. Ribosomi i Golgijev kompleks također su odsutni, ali je endoplazmatski retikulum dobro razvijen, koji ne samo da prodire u citoplazmu, već također prolazi u susjedne stanice kroz pore sitastih polja. Dobro razvijeni mitohondriji i plastidi nalaze se u izobilju.
Između stanica, tvari se transportiraju kroz rupe koje se nalaze na staničnim membranama. Takvi otvori se nazivaju pore, ali za razliku od pora traheida, oni su prolazni. Pretpostavlja se da predstavljaju jako proširene plazmodezme, na stijenkama, koje? taloži se polisaharid kaloza. Pore su raspoređene u skupine, tvoreći sitasta polja. U primitivnim oblicima polja sita nasumično su razbacana po cijeloj površini ljuske; u naprednijim angiospermama nalaze se na krajevima susjednih stanica jedna uz drugu, tvoreći sitastu ploču. Ako je na njemu jedno polje sita, naziva se jednostavnim, ako ih je više, naziva se složenim.
Brzina kretanja otopina kroz sitaste elemente je do 150 cm na sat. To je tisuću puta brže od brzine slobodne difuzije. Vjerojatno se odvija aktivni transport, a brojni mitohondriji sitastih elemenata i pratećih stanica opskrbljuju potreban ATP za to.
Trajanje aktivnosti sitastih elemenata floema ovisi o prisutnosti bočnih meristema. Ako su prisutni, tada elementi sita rade tijekom cijelog životnog vijeka biljke.
Osim sitastih elemenata i pratećih stanica, floem sadrži lična vlakna, sklereide i parenhim.
U procesu evolucije, izlaskom viših biljaka na kopno, razvile su se tkiva koje su najveću specijalizaciju postigle u cvjetnicama. U ovom članku ćemo pobliže pogledati što su biljna tkiva, koje vrste postoje, koje funkcije obavljaju, kao i strukturne značajke biljnih tkiva.
Tkanina su skupine stanica koje su slične strukture i obavljaju iste funkcije.
Glavna biljna tkiva prikazana su na donjoj slici:
Vrste, funkcije i građa biljnih tkiva.
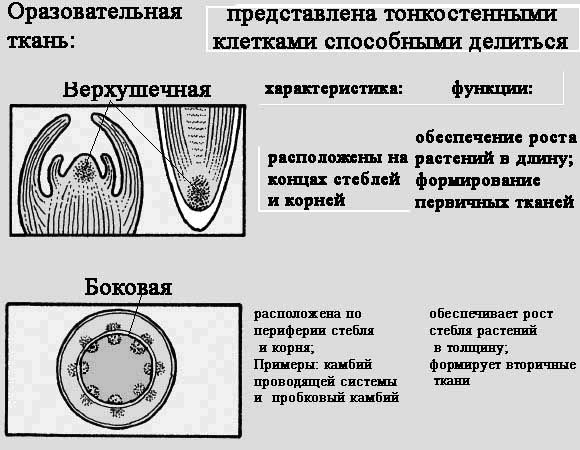

Pokrovno tkivo biljaka.


Biljno pokrovno tkivo – kora
Provodno biljno tkivo.
| Naziv tkanine | Struktura | Mjesto | Funkcije |
| 1. Drvene posude - ksilem | Šuplje cijevi s lignificiranim stijenkama i mrtvim sadržajem | Drvo (ksilem) koje se proteže duž korijena, stabljike, lisnih žila | Provođenje vode i minerala iz tla do korijena, stabljike, lišća, cvijeća |
|
2. Sitaste cijevi od lika - floema Popratne stanice ili prateće stanice |
Okomiti niz živih stanica sa sitastim poprečnim pregradama Sestrinske stanice sitastih elemenata koje su zadržale svoju strukturu |
Bast (floem), smješten duž korijena, stabljike, lisnih vena Uvijek se nalaze uz elemente sita (prate ih) |
Prenošenje organske tvari od lišća do stabljike, korijena, cvijeća Aktivno sudjelujte u prijenosu organskih tvari kroz sitaste cijevi floema |
| 3. Provođenje vaskularno-vlaknastih snopova | Kompleks drva i lišća u obliku zasebnih niti kod trava i kontinuirane mase kod drveća | Središnji cilindar korijena i stabljike; žile lišća i cvjetova | Provođenje vode i minerala kroz drvo; na bastu - organske tvari; jačanje organa, povezujući ih u jednu cjelinu |



Mehanička tkiva biljaka.

