Коріння та провідні тканини є у. Будова рослинної клітини
ПРОВОДНІ ТКАНИНИ.
Провідні тканини служать для пересування рослиною розчинених у воді поживних речовин.
Мал. 43 Дерев'яні волокна листа герані лугової (поперечний - А, Б і поздовжній - У розріз групи волокон):
1 - стінка клітини; 2 - прості пори; 3 - порожнина клітини.
Подібно до покривних тканин, вони виникли як наслідок пристосування рослини до життя у двох середовищах: ґрунтовому та повітряному. У зв'язку з цим виникла потреба транспортування поживних речовин у двох напрямках.
Від кореня до листя рухається висхідний, або транспіраційний струм водних розчинів солей. Асиміляційний, низхідний, струм органічних речовин прямує від листя до коріння. Висхідний струм здійснюється майже виключно по трахеальних

Мал. 44 Склереїди кісточки дозріваючих плодів аличі з живим вмістом: 1 - цитоплазма, 2 - потовщена клітинна оболонка, 3-порові канальці
елементам ксилеми, а. низхідний - за ситоподібними елементами флоеми.
Сильно розгалужена мережа провідних тканин несе водорозчинні речовини та продукти фотосинтезу до всіх органів рослини, починаючи від найтонших кореневих закінчень до наймолодших пагонів. Провідні тканини поєднують усі органи рослини. Крім далекого, тобто осьового, транспорту поживних речовин, по провідних тканинах здійснюється-і ближній - радіальний транспорт.
Усі провідні тканини є складними, або комплексними, тобто складаються з морфологічно та функціонально різнорідних елементів. Формуючись з однієї меристеми, два типи провідних тканин - ксилема і флоема - розташовуються поруч. У багатьох органах рослин ксилема поєднана з "флоемою у вигляді тяжів, званих провідними пучками.
Існують первинні та вторинні провідні тканини. Первинні тканини закладаються в листі, молодих пагонах і коренях. Вони диференціюються з клітин прокамбію. Вторинні провідні тканини, зазвичай потужніші, виникають із камбію.
Ксилема (деревина).По ксилемі від кореня до листя пересуваються вода і розчинені у ній мінеральні речовини. Первинна і вторинна ксилеми містять клітини тих самих типів. Однак первинна ксилема не має серцевинних променів, відрізняючись цим від вторинної.
До складу ксилеми входять морфологічно різні елементи, що здійснюють функції проведення, так і зберігання запасних речовин, а також суто опорні функції. Далекий транспорт здійснюється за трахеальними елементами ксілеми: трахеїдами та судинами, ближній - за паренхімними елементами. Опорні, а іноді й запасні функції виконують частину трахеїд і волокна механічної тканини лібриформу, що також входять до складу ксилеми.
Трахеїди у зрілому стані – це мертві прозенхімні клітини, звужені на кінцях та позбавлені протопласту. Довжина трахеїд в середньому становить 1-4 мм, діаметр же не перевищує десятих і навіть сотих часток міліметра. Стінки трахеїд дерев'яні, потовщуються і несуть прості або облямовані пори, через які відбувається фільтрація розчинів. Більшість облямованих пір знаходиться біля закінчень клітин, тобто там, де розчини просочуються з однієї трахеїди в іншу. Трахеїди є у спорофітів усіх вищих рослин, а у більшості хвощевидних, плауновидних, папоротеподібних і голонасінних вони є єдиними провідними елементами ксілеми.
Судини - це порожнисті трубки, що складаються з окремих члеників, що розташовуються одна над одною.
Між розташованими один над одним члениками однієї і тієї ж судини є різного типунаскрізні отвори – перфорації. Завдяки перфораціям вздовж усієї судини вільно здійснюється струм рідини. Еволюційно судини, мабуть, походять з трахеїд шляхом руйнування замикаючих плівок пір і подальшого їх злиття в одну або кілька перфорацій. Кінці трахеїд, спочатку сильно скошені, зайняли горизонтальне положення, а самі трахеїди стали коротшими і перетворилися на членики судин (рис. 45).
Судини з'явилися незалежно у різних лініях еволюції наземних рослин. Однак найбільшого розвитку вони досягають у покритонасінних, де є найголовнішими водопровідними елементами ксилеми. Виникнення судин – важливе свідчення еволюційного прогресу цього таксону, оскільки вони суттєво полегшують транспіраційний струм вздовж тіла рослини.
Крім первинної оболонки, судини та трахеїди в більшості випадків мають вторинні потовщення. У наймолодших трахеальних елементах вторинна оболонка може мати форму кілець, не пов'язаних один з одним (кільчасті трахеїди та судини). Пізніше виникають трахеальні елементи зі спіральними потовщеннями. 3$тем слідують судини та трахеїди з потовщеннями, які можуть бути охарактеризовані як спіралі, витки яких пов'язані між собою (сходові потовщення). Зрештою вторинна оболонка зливається у більш менш суцільний циліндр, що формується всередину від первинної оболонки. Цей циліндр переривається в окремих ділянках порами. Судини і трахеїди з відносно невеликими округлими ділянками первинної клітинної оболонки, не прикритими зсередини вторинною оболонкою, нерідко називають пористими. ).

Мал. 45 Зміна структури трахеальних елементів ксилеми в ході їх еволюції (напрямок позначений стрілкою):
1,2 - трахеїди з округлими облямованими порами, 3 - трахеїди з витягнутими облямованими порами, 4 - членик судини примітивного типу та його перфорація, утворена зближеними порами, 5 - 7 - послідовні стадії спеціалізації члеників судин та утворення простої перфорації
Вторинна, котрий іноді первинна оболонка, зазвичай, лигнифицируются, т. е. просочуються лігніном, це надає додаткову міцність, але обмежує можливості їх подальшого зростання довжину.
Трахеальні елементи, тобто трахеїди та судини, розподіляються в ксілемі по-різному. Іноді на поперечному зрізі вони утворюють добре виражені кільця (кільцесосудиста деревина). В інших випадках судини розсіяні більш менш рівномірно по всій масі ксілеми (розсіяносудинна деревина). Особливості розподілу трахеальних елементів у ксілемі використовують щодо деревини різних порід дерев.
Крім трахеальних елементів, ксилема включає променеві елементи, тобто клітини, що утворюють серцевинні промені (рис. 46), сформовані найчастіше тонкостінними паренхімними клітинами (променева паренхіма). Рідше в хвойних променях зустрічаються променеві трахеїди. По серцевим променям здійснюється ближній транспорт речовин у горизонтальному напрямку. У ксилемі покритонасінних, крім провідних елементів, містяться також тонкостінні неодревеснілі живі паренхімні клітини, які називаються деревинною паренхімою. По них поруч із серцевинними променями частково здійснюється ближній транспорт. Крім того, дерев'яна паренхіма є місцем зберігання запасних речовин. Елементи
серцевинних променів та деревинної паренхіми, подібно до трахеальних елементів, виникають з камбію.
Провідні тканини є складними, оскільки вони складаються з кількох типів клітин, їх структури мають витягнуту (трубчасту) форму, пронизані численними порами. Наявність отворів на торцевих (нижніх чи верхніх) ділянках забезпечують вертикальний транспорт, а пори на бічних поверхнях сприяє надходженню води у радіальному напрямку. До провідних тканин відносять ксилему та флоему. Вони є тільки у папоротеподібних та насіннєвих рослин. У провідній тканині є як мертві, так і живі клітини
Ксилема (деревина)- Це мертва тканина. Включає основні структурні компоненти (трахеї і трахеїди), деревинну паренхіму і дерев'яні волокна. Вона виконує в рослині як опорну, так і функцію, що проводить - по ній рухаються вгору по рослині вода і мінеральні солі.
Трахеїди
– мертві поодинокі клітини веретеноподібної форми. Стінки сильно потовщені внаслідок відкладення лігніну. Особливістю трахеїд є наявність у їх стінках облямованих пір. Їхні кінці перекриваються, надаючи рослині необхідну міцність. Вода рухається порожніми просвітами трахеїд, не зустрічаючи на своєму шляху перешкод у вигляді клітинного вмісту; від однієї трахеїди до іншої вона передається через пори.
У покритонасінних трахеїди розвинулися в судини (трахеї). Це дуже довгі трубки, що утворилися в результаті стикування ряду клітин; залишки торцевих перегородок все ще зберігаються в судинах у вигляді обідків-перфорацій. Розміри судин варіюють від кількох сантиметрів до кількох метрів. У перших за часом утворення судин протоксилеми лігнін накопичується кільцями або спіраллю. Це дає можливість посудині продовжувати розтягуватися під час зростання. У судинах метаксилеми лігнін зосереджений щільніше – це ідеальний «водопровід», що діє великі відстані.
?1.
Чим трахеї відрізняються від трахеїдів? (Відповідь наприкінці статті)
?2
. Чим трахеїди відрізняються від волокон?
?3
. Що спільного у флоеми та ксилеми?
?4.
Чим ситоподібні трубки відрізняються від трахей?
Паренхімні клітини ксілеми утворюють своєрідні промені, що з'єднують серцевину з корою. Вони проводять воду у радіальному напрямку, запасають поживні речовини. З інших клітин паренхіми розвиваються нові судини ксилеми. Нарешті, деревні волокна схожі на трахеїди, але на відміну від неї мають дуже малий внутрішній просвіт, тому не проводять воду, але надають додаткову міцність. А також мають прості пори, а не облямовані.
Флоема (луб)- Це жива тканина, що входить до складу кори рослин, по ній здійснюється низхідний струмводи з розчиненими у ній продуктами асиміляції. Флоема утворена п'ятьма типами структур: ситоподібні трубки, клітини-супутниці, луб'яна паренхіма, луб'яні волокна та склереїди.
Основою цих структур є ситоподібні трубки
, що утворюються в результаті з'єднання ряду ситоподібних клітин Їхні стінки тонкі, целюлозні, ядра після дозрівання відмирають, а цитоплазма притискається до стінок, звільняючи шлях для органічних речовин. Торцеві стінки клітин ситоподібних трубок поступово покриваються порами і починають нагадувати сито – це ситоподібні платівки. Задля більшої їх життєдіяльності поруч розташовуються клітини-супутниці, їх цитоплазма активна, ядра великі.
?5
. Як ви вважаєте, чому при дозріванні ситовидних клітин їхні ядра відмирають?
ВІДПОВІДІ
?1.
Трахеї багатоклітинні структури та торцевих стінок не мають, а трахеїди одноклітинні, мають торцеві стінки та облямовані пори.
?2
. Трахеїди облямовані пори та добре виражений просвіт, а у волокон просвіт дуже маленький і пори прості. Вони також відрізняються функціями, трахеїди виконують транспортну роль (що проводить), а волокна механічну.
?3.
Флоема і ксилема – обидві провідні тканини, їх структури мають трубчасту форму, до їх складу входять клітини паренхіми та механічних тканин.
?4.
Ситоподібні трубки складаються з живих клітин, їх целюлозні стінки, здійснюють низхідний транспорт органічних речовин, а трахеї утворені мертвими клітинами їх стінки сильно потовщені лігніном, забезпечують висхідний транспорт води і мінеральних речовин.
?5.
По ситоподібних клітин відбувається низхідний транспорт і ядра, що захоплюються струмом речовин, закривали б значну частину стоподібного поля що призводило б зниження ефективності процесу.
Провідні тканини служать для пересування рослиною розчинених у воді поживних речовин. Вони виникли як наслідок пристосування рослин до життя на суші. У зв'язку з життям у двох середовищах – ґрунтовому та повітряному, виникли дві провідні тканини, за якими речовини пересуваються у двох напрямках.
По ксилемі від коріння до листя піднімаються речовини ґрунтового харчування– вода та розчинені в ній мінеральні солі (висхідний, або транспіраційний струм).
По флоем від листя до коренів пересуваються речовини, що утворилися в процесі фотосинтезу, головним чином сахароза (низхідний струм). Так як ці речовини є продуктами асиміляції вуглекислого газу, транспорт речовин по флоемі називають струмом асимілятів.
Провідні тканини утворюють у тілі рослини безперервну розгалужену систему, що з'єднує всі органи - від найтонших корінців до наймолодших пагонів. Ксилема і флоема є складні тканини, До їх складу входять різнорідні елементи - провідні, механічні, запасні, видільні. Найважливішими є елементи, що проводять, саме вони виконують функцію проведення речовин.
Ксилема і флоема формуються з однієї і тієї ж меристеми і, тому, в рослині завжди розташовуються поряд. Вторинні провідні тканини мають складнішу будову, ніж первинні.
Ксилема (деревина)складається з провідних елементів - трахеїд та судин (трахей), механічних елементів -деревинних волокон (волокон лібриформа) та елементів основної тканини - деревинної паренхіми.
Провідні елементи ксилеми звуться трахеальних елементів. Розрізняють два типи трахеальних елементів – трахеїди та членики судин (рис. 3.26).
Трахеїда є сильно витягнутою в довжину клітиною з непорушеними первинними стінками. Пересування розчинів відбувається шляхом фільтрації через облямовані пори. Судина складається з багатьох клітин, званих члениками суду. Членики розташовані один над одним, утворюючи трубочку. Між сусідніми члениками однієї і тієї ж судини є наскрізні отвори – перфорації. По судинах розчини пересуваються значно легше, ніж по трахеїдах.
Мал. 3.26. Схема будови та поєднання трахеїд (1) та члеників судини (2).
Трахеальні елементи в зрілому стані, що функціонує, - мертві клітини, що не мають протопластів. Збереження протопластів ускладнювало б пересування розчинів.
Судини і трахеїди передають розчини у вертикальному, а й у горизонтальному напрямі у сусідні трахеальні елементи й у живі клітини. Бічні стінки трахеїд та судин зберігаються тонкими на більшій чи меншій площі. У той же час вони мають вторинні потовщення, що надають стінам міцності. Залежно від характеру потовщень бічних стінок трахеальні елементи називаються кільчастими, спіральними, сітчастими, сходовими і крапково-поровими (рис. 3.27).

Мал. 3.27. Типи потовщення та поровності бічних стінок у трахеальних елементів: 1 – кільчасте, 2-4 – спіральні, 5 – сітчасте потовщення; 6 – сходова, 7 – супротивна, 8 – чергова поровість.
Вторинні кільчасті та спіральні потовщення прикріплюються до тонкої первинної стінки у вигляді вузького виступу. При зближенні потовщень та освіті між ними перемичок виникає сітчасте потовщення, що переходить в облямовані пори. Цю серію (рис. 3.27) можна як морфогенетичний, еволюційний ряд.
Вторинні потовщення клітинних стінок трахеальних елементів здерев'янюють (просочуються лігніном), що надає їм додаткової міцності, але обмежує можливості зростання в довжину. Тому в онтогенезі органу спочатку з'являються ще здатні розтягуватися кільчасті та спіральні елементи, що не перешкоджають зростанню органу в довжину. Коли ріст органу припиняється, виникають елементи, нездатні до поздовжнього розтягування.
У процесі еволюції першими з'явилися трахеїди. Вони знайдені у перших примітивних наземних рослин. Судини з'явилися значно пізніше шляхом перетворення трахеїдів. Судинами володіють майже всі покритонасінні рослини. Спорові і голонасінні рослини, як правило, позбавлені судин і мають лише трахеїди. Лише у вигляді рідкісного виключення судини зустрінуті у таких спорових, як селагінелла, деяких хвощів і папоротей, а також у небагатьох голонасінних (гнітових). Однак у цих рослин судини виникли незалежно від судин покритонасінних. Виникнення судин у покритонасінних рослин означало важливе еволюційне досягнення, оскільки полегшило проведення води; покритонасінні рослини виявилися більш пристосованими до життя на суші.
Деревина паренхіма і деревні волокна виконують запасні і опорні функції відповідно.
Флоема (луб)складається з провідних - ситовидних - елементів, що супроводжують клітин (клітин-супутниць), механічних елементів – флоемних (луб'яних) волокон та елементів основної тканини – флоемної (луб'яної) паренхіми.
На відміну від трахеальних елементів провідні елементи флоеми й у зрілому стані залишаються живими, які клітинні стінки – первинними, неодревесневшими. На стінках ситоподібних елементівє групи дрібних наскрізних отворів – ситоподібні поля, якими повідомляються протопласти сусідніх клітин і відбувається транспорт речовин. Розрізняють два типи ситоподібних елементів – ситоподібні клітини та членики ситоподібних трубок.
Ситоподібні клітини є примітивнішими, вони притаманні споровим і голонасінним рослинам. Ситовидна клітина - це одна клітина, сильно витягнута в довжину, із загостреними кінцями. Її ситоподібні поля розсіяні з бокових стінок. Крім того, ситоподібні клітини мають інші примітивні ознаки: вони позбавлені спеціалізованих супроводжуючих клітин і в зрілому стані містять ядра.
У покритонасінних рослин транспорт асимілятів здійснюють ситоподібні трубки (рис. 3.28). Вони складаються з багатьох окремих клітин – члеників, які розташовані одна над одною. Ситоподібні поля двох сусідніх члеників утворюють сітоподібну платівку. Ситоподібні пластинки мають більш досконалу будову, ніж ситоподібні поля (перфорації більші та їх більше).
У члениках ситовидних трубок у зрілому стані відсутні ядра, проте вони залишаються живими та активно проводять речовини. Важлива роль у проведенні асимілятів з ситоподібним трубкамналежить супроводжуючим клітин (клітин-супутницям). Кожен членик ситовидної трубки і його клітина, що супроводжує (або дві-три клітини у разі додаткового поділу) виникають одночасно з однієї меристематичної клітини. Клітини-супутниці мають ядра та цитоплазму з численними мітохондріями; у них відбувається інтенсивний обмін речовин. Між ситоподібними трубками і прилеглими до них клітинами, що супроводжують, є численні цитоплазматичні зв'язки. Вважається, що клітини-супутниці разом із члениками ситовидних трубок становлять єдину фізіологічну систему, яка здійснює струм асимілятів.

Мал. 3.28. Флоема стебла гарбуза на поздовжньому (А) та поперечному (Б) зрізі: 1 – членик ситовидної трубки; 2 – ситоподібна платівка; 3 – клітина, що супроводжує; 4 – луб'яна (флоемна) паренхіма; 5 – закупорена ситоподібна платівка.
Тривалість функціонування ситоподібних трубок невелика. У однорічників та в надземних втечах багаторічних трав– не більше одного вегетаційного періоду, у чагарників та дерев – не більше трьох-чотирьох років. При відмиранні живого вмісту ситовидної трубки відмирає і клітина-супутниця.
Луб'яна паренхіма складається з живих тонкостінних клітин. У її клітинах часто накопичуються запасні речовини, і навіть смоли, танниды та інших. Луб'яні волокна грають опорну роль. Вони є не у всіх рослин.
У тілі рослини ксилема та флоема розташовані поруч, утворюючи або шари, або відокремлені тяжі, які називають провідними пучками. Розрізняють кілька типів пучків, що проводять (рис. 3.29).
Закриті пучки складаються тільки з первинних провідних тканин, вони не мають камбію і далі не потовщуються. Закриті пучки характерні для спорових та однодольних рослин. Відкриті пучки мають камбій та здатні до вторинного потовщення. Вони характерні для голонасінних та дводольних рослин.
Залежно від взаємного розташування флоеми та ксилеми в пучку розрізняють такі типи. Найбільш звичайні колатеральні пучки, в яких флоема лежить по один бік від ксилеми. Колатеральні пучки можуть бути відкритими (стебла дводольних та голонасінних рослин) та закритими (стебла однодольних рослин). Якщо з внутрішньої сторонивід ксилеми розташовується додатково тяж флоеми, такий пучок називається біколатеральним. Біколатеральні пучки можуть бути тільки відкритими, вони характерні для деяких сімейств дводольних рослин (гарбузові, пасльонові та ін.).
Зустрічаються також концентричні пучки, у яких одна провідна тканина оточує іншу. Вони можуть бути лише закритими. Якщо в центрі пучка знаходиться флоема, а ксилема її оточує, пучок називається центрофлоемним, або амфівазальним. Такі пучки часто зустрічаються у стеблах та кореневищах однодольних рослин. Якщо в центрі пучка розташовується ксилема і її оточує флоема, пучок називається центроксилемним, або амфікрибральним. Центроксилемні пучки звичайні у папороті.

Мал. 3.29. Типи пучків, що проводять: 1 – відкритий колатеральний; 2 – відкритий біколатеральний; 3 – закритий колатеральний; 4 – концентричний закритий центрофлоемний; 5 – концентричний закритий центроксілемний; К – камбій; Кс – ксилема; Ф – флоема.
5.Механічна, запасна, повітроносна тканини. Будова, функції
Механічна тканинавид тканини в рослинному організмі, волокна з живих і мертвих клітин з сильно потовщеною клітинною стінкою, що надають механічної міцності організму. Виникає з верхівкової меристеми, а також у результаті діяльності прокамбію та камбію.
Ступінь розвитку механічних тканин багато в чому залежить від умов, вони майже відсутні у рослин вологих лісів, у багатьох прибережних рослин, зате добре розвинені у більшості рослин посушливих місцепроживання.
Механічні тканиниприсутні у всіх органах рослини, але найбільше вони розвинені по периферії стебла і в центральній частині кореня.
Вирізняють такі типи механічних тканин:
Коленхіма - еластична опорна тканина первинної кори молодих стебел дводольних рослин, а також листя. Складається з живих клітин з нерівномірно потовщеними первинними оболонками, що не одеревіли, витягнутими вздовж осі органу. Створює опору рослині.
склеренхіма - міцна тканиназ швидко відмираючих клітин з здерев'янілими і рівномірно потовщеними оболонками. Забезпечує міцність органів та всього тіла рослин. Розрізняють два типи склеренхімних клітин:
волокна - довгі тонкі клітини, зазвичай зібрані в тяжі або пучки (наприклад, луб'яні або древні волокна).
склереїди - округлі мертві клітини з дуже товстими здерев'янілими оболонками. Ними утворені насіннєва шкірка, шкаралупа горіхів, кісточки вишні, сливи, абрикоса; вони надають м'якоті груш характерного крупчастого характеру. Зустрічаються групами у кірці хвойних та деяких листяних порід, у твердих оболонках насіння та плодів. Їхні клітини круглої формиз товстими стінками та маленьким ядром.
Механічні тканини забезпечують міцність органів рослин. Вони складають каркас, який підтримує всі органи рослин, протидіючи їх зламу, стиску, розриву. Основними характеристиками будови механічних тканин, що забезпечують їхню міцність і пружність, є потужне потовщення та здеревнення їх оболонок, тісне змикання між клітинами, відсутність перфорацій у клітинних стінках.
Механічні тканини найбільш розвинені в стеблі, де вони представлені луб'яними та деревними волокнами. У коренях механічна тканина зосереджена у центрі органу.
Залежно від форми клітин, їх будови, фізіологічного стану та способу потовщення клітинних оболонок розрізняють два види механічної тканини: коленхіму та склеренхіму, (рис. 8.4).

Мал. 8.4. Механічні тканини: а-кутова коленхіма; 6 склеренхіма; в - склереїди з плодів аличі: 1-цитоплазма, 2-потовщена клітинна стінка, 3-порові канальці.
Колленхіма представлена живими паренхімними клітинами з нерівномірно потовщеними оболонками, що роблять їх особливо добре пристосованими для зміцнення молодих органів. Будучи первинними, клітини коленхіми легко розтягуються і мало заважають подовженню тієї частини рослини, у якій перебувають. Зазвичай коленхіма розташовується окремими тяжами або безперервним циліндром під епідермою молодого стебла і черешків листя, а також облямовує жилки в дводольних листках. Іноді коленхіма містить хлоропласти.
Склеренхіма складається з витягнутих клітин з рівномірно потовщеними, часто здерев'янілими оболонками, вміст яких відмирає на ранніх стадіях. Оболонки склеренхімних клітин мають високу міцність, близьку до міцності сталі. Ця тканина широко представлена у вегетативних органах наземних рослин і становить їхню осьову опору.
Розрізняють два типи склеренхімних клітин: волокна та склереїди. Волокна - це довгі тонкі клітини, зазвичай зібрані в тяжі або пучки (наприклад, луб'яні або волокна деревини). Склереїди - це округлі мертві клітини з дуже товстими здерев'янілими оболонками. Ними утворені насіннєва шкірка, шкаралупа горіхів, кісточки вишні, сливи, абрикоси; вони надають м'якоті груш характерного крупчастого характеру.
Основна тканина, або паренхіма, складається з живих, зазвичай тонкостінних клітин, які складають основу органів (звідки і назва тканини). У ній розміщені механічні, провідні та інші постійні тканини. Основна тканина виконує ряд функцій, у зв'язку з чим розрізняють асиміляційну (хлоренхіму), запасну, повітроносну (аеренхіму) та водоносну паренхіму (рис. 8.5).

Рис. 8.5. Паренхімні тканини: 1-3 - хлорофілоносна (стовпчаста, губчаста та складчаста відповідно); 4-запасаюча (клітини із зернами крохмалю); 5 - повітроносна, або аеренхіма.
У клітинах запасає паренхіми відкладаються білки, вуглеводи та інші речовини. Вона добре розвинена у стеблах деревних рослин, у коренеплодах, бульбах, цибулинах, плодах та насінні. У рослин пустельних місцепроживання (кактуси) і солончаків у стеблах і листі є водоносна паренхіма, що служить для накопичення води (наприклад, у великих екземплярів кактусів з роду карнегія в тканинах міститься до 2-3 тис. л води). У водних і болотяних рослин розвивається особливий тип основної тканини - повітроносна паренхіма, або аеренхіма. Клітини аеренхіми утворюють великі повітроносні міжклітини, якими повітря доставляється до частин рослини, зв'язок яких із атмосферою утруднена.
Аеренхіма (або Еренхіма) - повітроносна тканина у рослин, побудована з клітин, з'єднаних між собою так, що між ними залишаються великі заповнені повітрям порожнечі (великі міжклітини).
У деяких посібниках аеренхіму розглядають як різновид основної паренхіми.
Побудована аеренхіма буває або зі звичайних паренхімних клітин, або з клітин зірчастої форми, з'єднаних один з одним своїми відрогами. Характеризується наявністю міжклітинників.
Зустрічається така повітроносна тканина у водних і болотних рослин, і призначення її подвійне. Насамперед це вмістище запасів повітря для потреб газового обміну. У рослин, повністю занурених у воду, умови газового обміну набагато менш зручні, ніж у наземних рослин. Тоді як останні оточені з усіх боків повітрям, водні рослини в кращому разі знаходять у навколишньому середовищідуже невеликі його запаси; ці запаси поглинаються вже поверхневими клітинами, а глибину товстих органів не доходять. Забезпечити собі нормальний газовий обмін рослина може за цих умов двома шляхами: або збільшуючи поверхню своїх органів при відповідному зменшенні їхньої масивності, або збираючи всередині своїх тканин запаси повітря. Обидва ці способи і спостерігаються насправді.
Газообмен.З одного боку, у багатьох рослин підводні листя надзвичайно сильно розсічені, як, наприклад, у водяного жовтця (англ.) рос. (Ranunculus aquatilis), Ouvirandrafene s tralis та ін.
З іншого боку, у разі масивності органів, вони є пухкою, наповненою повітрям губчастою масою. Протягом дня, коли, завдяки процесу асиміляції, рослина виділяє кисню в багато разів більше, ніж це необхідно для цілей дихання, виділений кисень і збирається про запас у великих міжклітинниках аеренхіми. У сонячну погодуЗначні кількості виділеного кисню не поміщаються у міжклітинниках і виходять назовні крізь різні випадкові отвори у тканинах. З настанням ночі, коли процес асиміляції припиняється, запасений кисень споживається поступово на дихання клітин, а натомість їх у повітроносні порожнини аеренхіми виділяється клітинами вуглекислота, щоб у свою чергу вдень піти на потреби асиміляції. Так вдень і вночі продукти життєдіяльності рослини, завдяки присутності аеренхіми, не витрачаються даремно, а залишаються про запас, щоб бути використаними в наступний період діяльності.
Що стосується рослин болотних, то в особливо невигідних умовах у сенсі дихання знаходяться у них коріння. Під шаром води, в просоченому водою грунті відбуваються різного роду процеси бродіння та гниття; кисень у самих верхніх шарахгрунту вже націло поглинений, далі створюються вже умови анаеробного життя, що протікає за відсутності кисню. Коріння болотних рослин не могли б існувати за таких умов, якби не було у них у розпорядженні запасу повітря в аеренхімі.
Відмінність болотних рослин і не цілком занурених водних рослинвід цілком занурених полягає в тому, що оновлення газів усередині аеренхіми відбувається не тільки завдяки життєдіяльності тканин, а й за допомогою дифузії (і термодифузії); в наземних органах система міжклітинників відкривається назовні масою дрібних отворів - продихів, крізь які шляхом дифузії і зрівнюється за складом повітря міжклітинників з навколишнім повітрям. Однак при дуже великих розмірахрослини такий шлях оновлення повітря в аеренхімі коренів був би недостатньо швидким. Відповідно до цього, наприклад, біля мангрових дерев, що ростуть по морських берегах з мулистим дном, деякі розгалуження коренів ростуть з мулу вгору і виносять у повітря над поверхнею води свої верхівки, поверхня яких пронизана численними отворами. Такі «дихальні корені» мають на меті більш швидке оновлення повітря в аеренхімі живильних коренів, розгалужених у безкисневому мулі морського дна.
Зменшення питомої ваги
Другим завданням аеренхіми є зменшення частки рослини. Тіло рослини важче за воду; аеренхіма грає для рослини роль плавального міхура; завдяки її присутності навіть тонкі, бідні механічними елементами органи тримаються у воді, а чи не падають безладно на дно. Підтримка органів, головним чином листя, у положенні, сприятливому для життєвих відправлень рослини, що досягається у наземних рослин дорогою ціною утворення маси механічних елементів, досягається тут у водяних рослин просто переповненням аеренхіми повітрям.
Особливо ясно виражена ця друга задача аеренхіми у плаваючого листя, де запити дихання могли б бути задоволені і без допомоги аеренхіми. Завдяки величезній кількості міжклітинних повітроносних ходів, лист не тільки плаває на поверхні води, та й здатний витримати деяку тяжкість. Особливо славляться цією властивістю гігантське листя Victoria regia. Аеренхіма, що виконує роль плавальних бульбашок, нерідко і утворює насправді на рослині міхурові здуття. Такі пухирі зустрічаються як у квіткових рослин (Eichhornia crassipes, Trianea bogotensis), так і у вищих водоростей: Sargassum bacciferum. Fucus vesiculosus та інші види забезпечені чудово розвиненими плавальними бульбашками.
У будь-якому живому чи рослинному організмі тканину утворюють подібні за походженням та будовою клітини. Будь-яка тканина пристосована до виконання однієї чи відразу кілька важливих для тваринного чи рослинного організму функцій.
Види тканин у вищих рослин
Виділяють такі види тканин рослин:
- освітні (меристема);
- покривні;
- механічні;
- провідні;
- основні;
- видільні.
Всі ці тканини мають свої особливості будови та відрізняються одна від одної виконуваними функціями.

Тканини рослин під мікроскопом
Освітня тканина рослин
Освітня тканина- Це первинна тканина, з якої утворюються всі інші тканини рослини. Вона складається з спеціальних клітин, здатних до багаторазового поділу. Саме з цих клітин складається зародок будь-якої рослини.
Ця тканина зберігається і у дорослої рослини. Вона розташовується:
ТОП-4 статтіякі читають разом з цією
- внизу кореневої системи та на верхівках стебел (забезпечує зростання рослини у висоту та розвиток кореневої системи) – верхівкова освітня тканина;
- всередині стебла (забезпечує зростання рослини завширшки, його потовщення) – бічна освітня тканина;
Покривна тканина рослин
Покривна тканинавідноситься до захисних тканин. Вона необхідна для того, щоб захищати рослину від різких перепадів температури, від зайвого випаровування води, від мікробів, грибів, тварин і від механічних пошкоджень.
Покривні тканини рослин утворені клітинами, живими та мертвими, здатними пропускати повітря, забезпечуючи необхідний зростання рослини газообмін.
Будова покривної тканини рослин така:
- спочатку розташована шкірка або епідерма, яка покриває листя рослини, стебла та найбільш уразливі частини квітки; клітини шкірки живі, еластичні, вони захищають рослину від зайвої втрати вологи;
- далі знаходиться пробка або перидерма, яка також розташовується на стеблах та коренях рослини (там, де утворюється шар пробки, шкірка відмирає); пробка захищає рослину від несприятливих впливів довкілля.
Також виділяють такий вид покривної тканини, як кірка. Ця міцна покривна тканина, пробка в даному випадку утворюється не тільки на поверхні, але і в глибині, причому верхні її шари потихеньку відмирають. По суті, кірка складається із пробки та мертвих тканин.

Рис.2 Корка – вид покривної тканини рослини
Для дихання рослини в кірці утворюються тріщини, на дні яких розташовуються спеціальні відростки, чечевички, через які відбувається газообмін.
Механічна тканина рослин
Механічні тканини надають рослині необхідну йому міцність. Саме завдяки наявності рослина може витримувати сильні пориви вітру і не ламаються під струменями дощу і під вагою плодів.
Виділяють два основні види механічних тканин: луб'яні та деревні волокна.
Проводять тканини рослин
Тканина, що проводить, забезпечує транспортування води з розчиненими в ній мінералами.
Ця тканина утворює дві транспортні системи:
- висхідну(від коріння до листя);
- низхідну(Від листя до всіх інших частин рослин).
Висхідна транспортна система складається з трахеїд і судин (ксилема або деревина), причому судини більш досконалі провідні засоби, ніж трахеїди.
У низхідних системах струм води з продуктами фотосинтезу проходить ситоподібними трубками (флоема або луб).
Ксилема та флоема утворюють судинно-волокнисті пучки – «кровоносну систему» рослини, яка пронизує її повністю, з'єднуючи в одне ціле.
Основна тканина
Основна тканина чи паренхіма– є основою рослини. У неї занурені всі інші види тканин. Це жива тканина і виконує різні функції. Саме через це виділяються різні її види (інформація про будову та функції різних видівосновна тканина представлена в таблиці нижче).
| Види основної тканини | Де знаходиться в рослині | Функції | Будова |
| Асиміляційна | листя та інші зелені частини рослини | сприяє синтезу органічних речовин | складається з фотосинтезуючих клітин |
| Запасаюча | бульби, плоди, нирки, насіння, цибулини, коренеплоди | сприяє накопиченню необхідних для розвитку рослини органічних речовин | тонкостінні клітини |
| Водоносна | стебло, листя | сприяє накопиченню води | пухка тканина, що складається з тонкостінних клітин |
| Повітроносна | стебло, листя, коріння | сприяє проведенню повітря по рослині | тонкостінні клітини |

Мал. 3 Основна тканина або паренхіма рослини
Видільні тканини
Назва цієї тканини свідчить, яку саме функцію вона грає. Ці тканини сприяють насиченню плодів рослин оліями та соками, а також сприяють виділенню листям, квітками та плодами особливого аромату. Таким чином, виділяють два види цієї тканини:
- тканини внутрішньої секреції;
- тканини зовнішньої секреції
Що ми дізналися?
Учням 6 класу до уроку біології слід запам'ятати, що тварини та рослини складаються з безлічі клітин, які, у свою чергу, упорядковано вишиковуються, утворюють ту чи іншу тканину. Ми з'ясували які види тканин існують у рослин – освітня, покривна, механічна, провідна, основна та видільна. Кожна тканина виконує свою, строго певну функцію, захищаючи рослину або забезпечуючи доступ усіх її частин до води чи повітря.
Тест на тему
Оцінка доповіді
Середня оцінка: 3.9. Усього отримано оцінок: 1552.
У процесі еволюції з виходом вищих рослин на сушу вони виникли тканини, які досягли своєї найбільшої спеціалізації у квіткових рослин. У цій статті ми розглянемо докладніше, що є тканини рослин, які види їх існують, які функції вони виконують, а також особливості будови тканин рослин.
Тканину називають групи клітин, подібних за своєю будовою та виконують однакові функції..
Основні тканини рослин представлені на малюнку нижче:
Види, функції та будова тканин рослин.


Покривна тканина рослин.
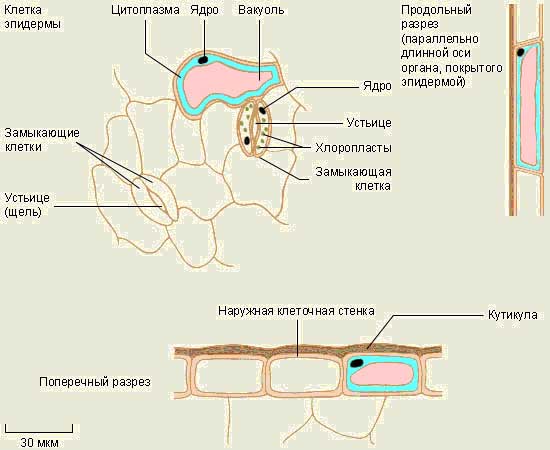


Покривна тканина рослин - кірка
Тканина рослин, що проводить.
| Назва тканини | Будова | Місцезнаходження | Функції |
| 1. Судини деревини – ксилема | Порожні трубки з стінами, що одеревіють, і відмерлим вмістом | Деревина (ксилема), що проходить уздовж кореня, стебла, жилок листя | Проведення води та мінеральних речовин із ґрунту в корінь, стебло, листя, квітки |
|
2.Ситовидні трубки лубу - флоема Супроводжуючі клітини або клітини-супутниці |
Вертикальний ряд живих клітин із ситоподібними поперечними перегородками Сестринські клітини ситоподібних елементів, що зберегли свою структуру |
Луб (флоема), розташований уздовж кореня, стебла, жилок листя Завжди розташовуються вздовж ситоподібних елементів (супроводжують їх) |
Проведення органічних речовин із листя в стебло, корінь, квітки Приймають активну участь у проведенні органічних речовин з ситоподібних трубок флоеми |
| 3. Провідні судинно-волокнисті пучки | Комплекс з деревини та лубу у вигляді окремих тяжів у трав та суцільного масиву у дерев | Центральний циліндр кореня та стебла; жилки листя та квіток | Проведення з деревини води та мінеральних речовин; по лубу – органічних речовин; зміцнення органів, зв'язок їх у єдине ціле |



Механічна тканина рослин.

