Функції провідних тканин квіткових рослин. Механічні та провідні тканини рослин
Тканина, що проводить, складається з живих або мертвих подовжених клітин, які мають вигляд трубок.
У стеблі та листі рослин розташовані пучки провідної тканини. У провідній тканині виділяють судини та ситоподібні трубки.
Судини- Послідовно з'єднані мертві порожнисті клітини, поперечні перегородки між якими зникають. По судинах вода і розчинені в ній мінеральні речовини з коренів надходять у стебло та листя.
Ситоподібні трубки - подовжені без'ядерні живі клітини, послідовно з'єднані між собою. По ним органічні речовини з листя (де вони утворилися) переміщаються до інших органів рослини.
Тканина, що проводить, забезпечує транспортування води з розчиненими в ній мінералами.
Ця тканина утворює дві транспортні системи:
- висхідну(від коріння до листя);
- низхідну(Від листя до всіх інших частин рослин).
Висхідна транспортна система складається з трахеїд і судин (ксилема або деревина), причому судини більш досконалі провідні засоби, ніж трахеїди.
У низхідних системах струм води з продуктами фотосинтезу проходить по ситоподібним трубкам(Флоема чи луб).
Ксилема та флоема утворюють судинно-волокнисті пучки – «кровоносну систему» рослини, яка пронизує її повністю, з'єднуючи в одне ціле.
Вчені вважають, що виникнення тканин пов'язане історія Землі з виходом рослин на сушу. Коли частина рослини опинилася в повітряному середовищі, а інша частина (коренева) – у ґрунті, з'явилася необхідність доставки води та мінеральних солей від коріння до листя, а органічних речовин – від листя до коріння. Так у ході еволюції рослинного світу виникло два типи провідних тканин – деревина та луб.
По деревині (по трахеїдах і судинах) вода з розчиненими мінеральними речовинами піднімається від коріння до листя - це водопровідний, або висхідний струм. По лубу (по ситоподібних трубках) органічні речовини, що утворилися в зеленому листі, надходять до коренів та інших органів рослини - це низхідний струм.
Освітня тканина
Освітня тканина знаходиться у всіх зростаючих частинах рослини. Освітня тканина складається з клітин, які здатні ділитися протягом усього життя рослини. Клітини тут лежать дуже швидко. Завдяки поділу вони утворюють безліч нових клітин, забезпечуючи цим зростання рослини в довжину і товщину. З'явилися під час поділу освітніх тканин клітини потім перетворюються на клітини інших тканин рослини.

Це первинна тканина, з якої утворюються й інші тканини рослини. Вона складається з спеціальних клітин, здатних до багаторазового поділу. Саме з цих клітин складається зародок будь-якої рослини.
Ця тканина зберігається і у дорослої рослини. Вона розташовується:
- внизу кореневої системи та на верхівках стебел (забезпечує зростання рослини у висоту та розвиток кореневої системи) – верхівкова освітня тканина;
- всередині стебла (забезпечує зростання рослини завширшки, його потовщення) – бічна освітня тканина.
На відміну від інших тканин, цитоплазма освітньої тканини густіша і щільніша. Клітина має добре розвинені органоїди, які забезпечують синтез білка. Ядру характерні великі розміри. Маса ядра та цитоплазми підтримуються у постійному співвідношенні. Збільшення ядра сигналізує початок процесу клітинного поділу, що відбувається шляхом мітозу для вегетативних частин рослин і мейозу для спорогенных меристем.
Значення та різноманітність провідних тканин
Провідні тканини є найважливішою складовою більшості вищих рослин. Вони є обов'язковим структурним компонентом вегетативних та репродуктивних органів спорових та насіннєвих рослин. Провідні тканини в сукупності з клітинними стінками і міжклітинниками, деякими клітинами основної паренхіми та спеціалізованими передатними клітинами утворюють провідну систему, яка забезпечує далекий та радіальний транспорт речовин. Завдяки особливій конструкції клітин та їх розташування в тілі рослин провідна система виконує численні, але взаємопов'язані функції:
1) пересування води та мінеральних речовин, поглинених корінням із ґрунту, а також органічних речовин, що утворюються в коренях, у стебло, листя, репродуктивні органи;
2) пересування продуктів фотосинтезу із зелених частин рослини у місця їх використання та запасання: у коріння, стебла, плоди та насіння;
3) пересування фітогормонів по рослині, що створює певний їх баланс, який визначає темпи зростання та розвитку вегетативних та репродуктивних органів рослин;
4) радіальний транспортречовин з провідних тканин у поруч розташовані живі клітини інших тканин, наприклад, в асимілюючі клітини мезофілу листа і клітини меристем, що діляться. У ньому можуть також брати участь паренхімні клітини серцевих променів деревини та кори. Велике значення в радіальному транспорті мають передавальні клітини з численними випинання клітинної оболонки, що знаходяться між провідними і паренхімними тканинами;
5) провідні тканини підвищують стійкість органів рослин до деформуючих навантажень;
6) провідні тканини утворюють безперервну розгалужену систему, яка зв'язує органи рослин в єдине ціле;
Виникнення провідних тканин є результатом еволюційних структурних перетворень, пов'язаних з виходом рослин на сушу та поділом їх повітряного та ґрунтового харчування. Найбільш древні провідні тканини – трахеїди виявлені у копалин рініофітів. Найвищого розвиткувони досягли у сучасних покритонасінних.
У процесі індивідуального розвитку первинні провідні тканини утворюються з прокамбію в точках зростання зародка насіння та нирок відновлення. Вторинні провідні тканини, характерні для дводольних покритонасінних, породжуються камбієм.
Залежно від виконуваних функцій провідні тканини поділяються на тканини висхідного струму та тканини низхідного струму. Основним призначенням тканин висхідного струму є транспорт води та розчинених у ній мінеральних речовин від кореня до вище розташованих надземних органів. Крім того, по них переміщуються органічні речовини, що утворюються в корені та стеблі, наприклад, органічні кислоти, вуглеводи та фітогормони. Однак термін "висхідний струм" не слід сприймати однозначно як пересування знизу - вгору. Тканини висхідного струму забезпечують потік речовин у напрямку від зони всмоктування до апексу втечі. При цьому речовини, що транспортуються, використовуються як самим коренем, так і стеблом, гілками, листям, репродуктивними органами, незалежно від того, вище або нижче рівня коренів вони знаходяться. Наприклад, у картоплі вода та елементи мінерального живлення надходять по тканинах висхідного струму в столони та бульби, що утворюються у ґрунті, а також у надземні органи.
Тканини низхідного струмузабезпечують відтік продуктів фотосинтезу в зростаючі частини рослин і запасні органи. У цьому просторове становище фотосинтезирующих органів немає значення. Наприклад, у пшениці органічні речовини надходять у зернівки, що розвиваються, з листя різних ярусів. Тому до назв «висхідні» і «низхідні» тканини слід відноситися не більше як до традиції, що склалася.
Провідні тканини висхідного струму
До тканин висхідного струму відносяться трахеїди та судини (трахеї), які розташовуються в деревинній (ксилемній) частині органів рослин. У цих тканинах пересування води та розчинених у ній речовин відбувається пасивно під дією кореневого тиску та випаровуванням води з поверхні рослини.
Трахеїди мають більш давнє походження. Вони зустрічаються у вищих спорових рослин, голонасінних і рідше – у покритонасінних. У покритонасінних вони типові для найдрібніших розгалужень жилок листа. Клітини трахеїд мертві. Вони мають витягнуту, часто веретеноподібну форму. Їхня довжина становить 1 – 4 мм. Однак у голонасінних, наприклад, у араукарії, вона досягає 10 мм. Стіни клітин товсті, целюлозні, часто просочуються лігніном. У клітинних оболонках є численні облямовані пори.
Судини сформувалися більш пізніх етапах еволюції. Вони характерні для покритонасінних, хоча зустрічаються і в деяких сучасних представників відділів Плауни (рід Селлагінелла), Хвощі, Папороті та Голосонасінні (рід Гнетум).
Судини складаються з подовжених мертвих клітин, розташованих одна над іншою і званих члениками судини. У торцевих стінках члеників судини є великі наскрізні отвори – перфорації, якими здійснюється далекий транспорт речовин. Перфорації виникли в ході еволюції з облямованих пір трахеїд. У складі судин вони бувають сходовими та простими. Численні сходові перфорації утворюються на торцевих стінках члеників судини за її косому закладенні. Отвори таких перфорацій мають подовжену форму, а перегородки, що їх розділяють, розташовуються паралельно один одному, нагадуючи сходи сходів. Судини зі сходовою перфорацією характерні для рослин сімейств Лютикові, Лимонникові, Березові, Пальмові, Частухові.
Прості перфорації відомі у еволюційно молодших сімейств, таких як Пасльонові, Гарбузові, Астрові, Мятлікові. Вони є одним великим отвіром в торцевій стінці членика, розташованої перпендикулярно осі судини. У ряді сімейств, наприклад, у Магнолієвих, Рожевих, Ірисових, Астрових, у судинах зустрічаються як прості, так і сходові перфорації.
Бічні стінки мають нерівномірні целюлозні потовщення, які оберігають судини від надлишкового тиску, що створюється поруч розташованими живими клітинами інших тканин. У бічних стінках можуть бути численні пори, які забезпечують вихід води межі судини.
Залежно від характеру потовщень, типів і характеру розташування пір судини поділяються на кільчасті, спіральні, біспіральні, сітчасті, сходові та точково-порові. У кільчастих та спіральних судин целюлозні потовщення розташовуються у вигляді кілець або спіралей. Через непотовщені ділянки здійснюється дифузія розчинів, що транспортуються в навколишні тканини. Діаметр цих судин порівняно невеликий. У сітчастих, сходових і точково-порових судин вся бічна стінка, за винятком місць розташування простого часу, потовщена і часто просочена лігніном. Тому радіальний транспорт речовин у них здійснюється через численні подовжені та точкові пори.
Судини мають обмежений термін. Вони можуть руйнуватися внаслідок закупорки тилами – виростами сусідніх паренхімних клітин, а також під дією доцентрових сил тиску нових клітин деревини, що утворюються камбієм. У ході еволюції судини зазнають змін. Членники судин стають коротшими і товстішими, косі поперечні перегородки змінюються прямими, а сходові перфорації - простими.
Провідні тканини низхідного струму
До тканин низхідного струму відносяться ситоподібні клітини та ситоподібні трубки з клітинами-супутницями. Ситоподібні клітини мають більш давнє походження. Вони зустрічаються у вищих спорових рослин та голонасінних. Це живі, видовжені клітини із загостреними кінцями. У зрілому стані вони містять ядра у складі протопласту. У їх бокових стінках, у місцях зіткнення суміжних клітин, є дрібні наскрізні перфорації, які зібрані групами і утворюють ситоподібні поля, якими здійснюється пересування речовин.
Ситоподібні трубки складаються з вертикального ряду подовжених клітин, розділених між собою поперечними стінками та званими ситоподібними пластинками, в яких розташовані ситоподібні поля. Якщо ситоподібна пластинка має одне ситоподібне поле, вона вважається простою, а якщо декількома – то складною. Ситоподібні поля утворюються численними наскрізними отворами – ситоподібними перфораціями невеликого діаметра. Через ці перфорації з однієї клітини до іншої проходять плазмодесми. На стінках перфорацій розміщується полісахарид калозу, що зменшує просвіт перфорацій. У міру старіння ситовидної трубки калоз повністю закупорює перфорації і трубка припиняє роботу.
При формуванні ситовидної трубки у клітинах, що їх утворюють, синтезується спеціальний флоемний білок (Ф-білок) і розвивається велика вакуоля. Вона відтісняє цитоплазму та ядро до стінки клітини. Потім мембрана вакуолі руйнується і внутрішній простірклітини заповнюється сумішшю цитоплазми та клітинного соку. Тільця Ф-білка втрачають чіткі контури, зливаються, утворюючи тяжи біля ситоподібних пластинок. Їхні фібрили проходять через перфорації з одного членика ситовидної трубки в інший. До члеників ситовидної трубки щільно прилягають одна або дві клітини-супутниці, які мають подовжену форму, тонкі стінки та живу цитоплазму з ядром та численними мітохондріями. У мітохондріях синтезується АТФ, необхідна для транспортування речовин ситоподібними трубками. У стінках клітин-супутниць є велика кількістьпір з плазмадесмами, яке майже в 10 разів перевищує їх кількість в інших клітинах мезофілу листа. Поверхня протопласту цих клітин значно збільшена за рахунок численних складок, що утворюються плазмалемою.
Швидкість пересування асимілятів ситоподібними трубками значно перевищує швидкість вільної дифузії речовин і досягає 50 - 150 см/год, що вказує на активний транспорт речовин з використанням енергії АТФ.
Тривалість роботи ситоподібних трубок у багаторічних дводольних становить 1 – 2 роки. На зміну їм камбій постійно утворює нові провідні елементи. У однодольних, позбавлених камбію, ситоподібні трубки є набагато довше.
Провідні пучки
Провідні тканини розташовуються в органах рослин у вигляді поздовжніх тяжів, утворюючи пучки, що проводять. Розрізняють чотири типи провідних пучків: прості, загальні, складні та судинно-волокнисті.
Прості пучки складаються з одного типу провідних тканин. Наприклад, у крайових частинах листових пластинок багатьох рослин зустрічаються невеликі по діаметру пучки з судин і трахеїд, а в квітконосних пагонах у лілейних - лише з ситовидних трубок.
Загальні пучки утворюються трахеїдами, судинами та ситоподібними трубками. Іноді цей термін використовується для позначення пучків метамеру, які проходять у міжвузлі та є листовими слідами. До складу складних пучків входять провідні та паренхімні тканини. Найбільш досконалими, різноманітними за будовою та місцем розташування є судинно-волокнисті пучки.
Судинно-волокнисті пучки характерні для багатьох вищих спорових рослин та голонасінних. Однак вони найбільш типові для покритонасінних. У таких пучках виділяються функціонально різні частини – флоема та ксилема. Флоема забезпечує відтік асимілятів з листа та пересування їх у місця використання чи запасання. По ксилемі вода та розчинені в ній речовини пересуваються з кореневої системи в лист та інші органи. Об'єм ксилемної частини в кілька разів перевищує обсяг флоемної, оскільки обсяг води, що надходить в рослину, перевищує обсяг утворюваних асимілятів, так як значна частина води випаровується рослиною.
Різноманітність судинно-волокнистих пучків визначається їх походженням, гістологічним складом та місцезнаходженням у рослині. Якщо пучки утворюються з прокамбію і завершують свій розвиток у міру використання запасу клітин освітньої тканини, як у однодольних, вони називаються закритими для зростання. На відміну від них, у дводольних відкриті пучки не обмежені у зростанні, оскільки вони формуються камбієм і збільшуються в діаметрі протягом усього життя рослини. До складу судинно-волокнистих пучків крім провідних можуть входити основні та механічні тканини. Наприклад, у дводольних флоема утворюється ситовидними трубками (провідна тканина висхідного струму), луб'яною паренхімою (основна тканина) і луб'яними волокнами (механічна тканина). До складу ксилеми входять судини та трахеїди (що проводить тканину низхідного струму), деревна паренхіма (основна тканина) та деревні волокна (механічна тканина). Гістологічний склад ксілеми та флоеми генетично детермінований і може бути використаний у систематиці рослин для діагностики різних таксонів. Крім того, ступінь розвитку складових частинпучків може змінюватися під впливом умов зростання рослин.
Відомо кілька видів судинно-волокнистих пучків.
Закриті колатеральні провідні пучки характерні для листя та стебел однодольних покритонасінних. Вони відсутні камбій. Флоема і ксилема розташовуються пліч-о-пліч. Для них характерні деякі конструктивні особливості. Так, у пшениці, що відрізняється С 3 шляхом фотосинтезу, пучки утворюються з прокамбію і мають первинну флоему і первинну ксилему. У флоемі виділяють більш ранню протофлоему і пізнішу за часом утворення, але більш великоклітинну метафлоему. У флоемній частині відсутні луб'яна паренхіма та луб'яні волокна. У ксілемі спочатку утворюються дрібніші судини протоксилеми, розташовані в одну лінію перпендикулярно до внутрішньої межі флоеми. Метаксилема представлена двома великими судинами, розташованими поруч із метафлоемою перпендикулярно ланцюжку судин протоксилеми. І тут судини розташовуються Т-образно. Відомо також V-, Y- та È-подібне розташування судин. Між судинами метаксилеми в 1 - 2 ряди розташована дрібноклітинна склеренхіма з потовщеними стінками, які в міру розвитку стебла просочуються лігніном. Ця склеренхіма відокремлює зону ксілеми від флоеми. По обидва боки від судин протоксилеми розташовуються клітини деревинної паренхіми, які, ймовірно, виконують трансфузійну роль, оскільки при переході пучка з міжвузля в листову подушку стеблового вузла вони беруть участь в утворенні клітин. Навколо пучка стебла пшениці, що проводить, розташовується склеренхімна обкладка, краще розвинена з боку протоксилеми і протофлоеми, біля бічних сторін пучка клітини обкладки розташовуються в один ряд.
У рослин з 4-типом фотосинтезу (кукурудза, просо та ін) в листі навколо закритих провідних пучків розташовується обкладка з великих клітин хлоренхіми.

Відкриті колатеральні пучки характерні для дводольних стебел. Наявність шару камбію між флоемою та ксилемою, а також відсутність склеренхімної обкладки навколо пучків забезпечує їх тривале зростання у товщину. У ксилемній та флоемній частинах таких пучків є клітини основної та механічної тканин.
Відкриті колатеральні пучки можуть бути утворені двома шляхами. По-перше, це пучки, які первинно утворюються прокамбієм. Потім у них із клітин основної паренхіми розвивається камбій, що виробляє вторинні елементи флоеми та ксилеми. В результаті пучки поєднують гістологічні елементи первинного та вторинного походження. Такі пучки характерні для багатьох трав'янистих квіткових рослин класу дводольних, що мають пучковий тип будови стебла (бобові, рожеві та ін.).
По-друге, відкриті колатеральні пучки можуть бути утворені лише камбієм і складатися з ксилеми та флоеми вторинного походження. Вони типові для трав'янистих дводольних з перехідним типом анатомічної будови стебла (астрові та ін), а також для коренеплодів типу буряків.
У стеблах рослин ряду сімейств (Гарбузові, Пасльонові, Дзвіночкові та ін) зустрічаються відкриті біколатеральні пучки, де ксилема з двох сторін оточена флоемою. При цьому зовнішня ділянка флоеми, звернена до поверхні стебла, розвинена краще внутрішнього, а смужка камбію, як правило, розташовується між ксилемою і зовнішньою ділянкою флоеми.
Концентричні пучки бувають двох типів. У амфікрибральних пучках, характерних для кореневищ папороті, флоема оточує ксилему, в амфівазальних – ксилема кільцем розташована навколо флоеми (кореневища ірису, конвалії та ін.). Рідше концентричні пучки зустрічаються у дводольних (кліщевина).
Закриті радіальні провідні пучки утворюються в ділянках коренів, що мають первинне анатомічна будова. Радіальний пучок входить до складу центрального циліндра та проходить через середину кореня. Його ксилема має вигляд багатопроменевої зірки. Між променями ксілеми розташовуються клітини флоеми. Число променів ксілеми значною мірою залежить від генетичної природи рослин. Наприклад, у моркви, буряків, капусти та інших дводольних ксилема радіального пучка має лише два промені. У яблуні та груші їх може бути 3 – 5, у гарбуза та бобів – ксилема чотирипроменева, а у однодольних – багатопроменева. Радіальне розташування променів ксілеми має пристосувальне значення. Воно скорочує шлях води від поверхні, що всмоктує, кореня до судин центрального циліндра.
У багаторічних деревних рослин і деяких трав'янистих однорічників, наприклад, у льону, провідні тканини розташовуються в стеблі, не утворюючи чітко виражених провідних пучків. Тоді говорять про непучковий тип будови стебла.
Тканини, що регулюють радіальний транспорт речовин
До специфічних тканин, що регулюють радіальний транспорт речовин відносяться екзодерми та ендодерма.

Екзодерма є зовнішнім шаром первинної кори кореня. Вона утворюється безпосередньо під первинною покривною тканиною епіблемою в зоні кореневих волосків і складається з одного або декількох шарів щільно зімкнутих клітин з потовщеними целюлозними оболонками. В екзодермі вода, що надійшла в корінь кореневими волосками, відчуває опір в'язкої цитоплазми і переміщається в целюлозні оболонки клітин екзодерми, а потім виходить з них в міжклітини середнього шару первинної кори, або мезодерми. Це забезпечує ефективне надходження води у глибші шари кореня.
У зоні проведення корені однодольних, де клітини епіблеми відмирають і злущуються, екзодерма виявляється на поверхні кореня. Її клітинні стінки просочуються суберином і перешкоджають надходженню води із ґрунту в корінь. У дводольних екзодерми у складі первинної кори злущується при линянні кореня і заміщається перидермою.
Ендодерма, або внутрішній шар первинної кори кореня, розташований навколо центрального циліндра. Вона утворюється одним шаром щільно зімкнутих клітин неоднакової будови. Одні з них, названі пропускними, мають тонкі оболонки і легко проникні для води. За ними вода з первинної кори надходить у радіальний провідний пучок кореня. Інші клітини мають специфічні целюлозні потовщення радіальних та внутрішніх тангентальних стінок. Ці потовщення, просочені суберином, називаються поясками Каспарі. Вони непроникні для води. Тому вода надходить у центральний циліндр лише через пропускні клітини. А оскільки поглинаюча поверхня кореня значно перевищує сумарну площу перерізу пропускних клітин ендодерми, то при цьому виникає кореневий тиск, який є одним із механізмів надходження води в стебло, лист та репродуктивні органи.
Ендодерма входить також до складу кори молодого стебла. У деяких трав'янистих покритонасінних вона, як і в корені, може мати пояски Каспарі. Крім того, в молодих стеблах ендодерма може бути представлена крохмалоносною піхвою. Таким чином, ендодерма може регулювати транспорт води у рослині та запасати поживні речовини.
Поняття про стелу та її еволюцію
Виникненню, розвитку в онтогенезі та еволюційним структурним перетворенням провідної системи приділяється велика увага, оскільки вона забезпечує взаємозв'язок органів рослин та з нею пов'язана еволюція великих таксонів.
На пропозицію французьких ботаніків Ф. Ван Тигема і А. Дуліо (1886) сукупність первинних провідних тканин разом із іншими тканинами і перициклом, що прилягають до кори, була названа стелою. До складу стели може також входити серцевина і утворена її місці порожнину, як, наприклад, у мятликових. Поняття "стела" відповідає поняттю "центральний циліндр". Стела кореня та стебла функціонально єдина. Вивчення стели у представників різних відділів вищих рослин призвело до формування стелярної теорії.
Розрізняють два основні типи стели: протостелу та еустелу. Найбільш давньою є протостела. Її провідні тканини розташовуються в середині осьових органів, причому в центрі знаходиться ксилема, оточена суцільним шаром флоеми. Серцевина чи порожнина у стеблі відсутні.
Існує кілька еволюційно пов'язаних між собою видів протостели: гаплостела, актиностела та плектостела.
Вихідним, примітивним виглядом є гаплостели. У неї ксилема має округлу форму поперечного перерізу і оточена безперервним рівним шаром флоеми. Навколо провідних тканин одним - двома шарами розташовується перицикл. Гаплостела була відома у копалин рініофітів і збереглася у деяких псилотофітів (тмезіптер).
Більше розвиненим видомпротостели є актиностела, в якій ксилема на поперечному перерізі набуває форми багатопроменевої зірки. Вона виявлена у викопного астероксилону та деяких примітивних плауноподібних.
Подальше роз'єднання ксилеми на окремі ділянки, розташовані радіально або паралельно один до одного, призвело до утворення плектостели, характерної для плауноподібних стебел. У актиностели та плектостели флоема, як і раніше, оточує ксилему з усіх боків.
У ході еволюції з протостели виникла сифоностела, відмінною особливістюякою є трубчаста будова. У центрі такої стели розташовується серцевина чи порожнина. У провідній частині сифоностели з'являються листові щілини, завдяки яким виникає безперервний зв'язок серцевини з корою. Залежно від способу взаємного розташування ксилеми і флоеми сифоностела буває ектофлою та амфіфлою. У першому випадку флоема з однієї, зовнішньої сторони оточує ксилему. У другому – флоема оточує ксилему із двох сторін, із зовнішньої та внутрішньої.
При поділі амфіфлойної сифоностели на мережу або ряди поздовжніх тяжів виникає розсічена стела, або диктіостела, характерна для багатьох папоротеподібних. Її провідна частина представлена численними концентричними пучками, що проводять.
У хвощів з ектофлойної сифоностели виникла артростела, яка має членисту будову. Вона відрізняється наявністю однієї великої центральної порожнини та відокремлених провідних пучків з протоксилемними порожнинами (каринальними каналами).
У квіткових рослин на основі ектофлойної сифоностели утворилася еустела, характерна для дводольних, і атактостела, типова для однодольних. В еустелі провідна частина складається з відокремлених колатеральних пучків, що мають кругове розташування. У центрі стели в стеблі розташовується серцевина, яка за допомогою серцевих променів з'єднується з корою. В атактостелі провідні пучки мають розсіяне розташування, між ними знаходяться паренхімні клітини центрального циліндра. Таке розташування пучків приховує трубчасту конструкціюсифоностели.
Виникнення різних варіантівсифоностели є важливим пристосуванням вищих рослин до збільшення діаметра осьових органів – кореня та стебла.
6.1. Значення та різноманітність провідних тканин
Провідні тканини є найважливішою складовою більшості вищих рослин. Вони є обов'язковим структурним компонентом вегетативних та репродуктивних органів спорових та насіннєвих рослин. Провідні тканини в сукупності з клітинними стінками і міжклітинниками, деякими клітинами основної паренхіми та спеціалізованими передатними клітинами утворюють провідну систему, яка забезпечує далекий та радіальний транспорт речовин. Завдяки особливій конструкції клітин та їх розташування в тілі рослин провідна система виконує численні, але взаємопов'язані функції:
1) пересування води та мінеральних речовин, поглинених корінням із ґрунту, а також органічних речовин, що утворюються в коренях, у стебло, листя, репродуктивні органи;
2) пересування продуктів фотосинтезу із зелених частин рослини у місця їх використання та запасання: у коріння, стебла, плоди та насіння;
3) пересування фітогормонів по рослині, що створює певний їх баланс, який визначає темпи зростання та розвитку вегетативних та репродуктивних органів рослин;
4) радіальний транспорт речовин з провідних тканин у поруч розташовані живі клітини інших тканин, наприклад, в асимілюючі клітини мезофілу листа і клітини меристем, що діляться. У ньому можуть також брати участь паренхімні клітини серцевих променів деревини та кори. Велике значення в радіальному транспорті мають передавальні клітини з численними випинання клітинної оболонки, що знаходяться між провідними і паренхімними тканинами;
5) провідні тканини підвищують стійкість органів рослин до деформуючих навантажень;
6) провідні тканини утворюють безперервну розгалужену систему, яка зв'язує органи рослин в єдине ціле;
Виникнення провідних тканин є результатом еволюційних структурних перетворень, пов'язаних з виходом рослин на сушу та поділом їх повітряного та ґрунтового живлення. Найбільш древні провідні тканини – трахеїди виявлені у копалин рініофітів. Найвищого розвитку вони досягли у сучасних покритонасінних.
У процесі індивідуального розвитку первинні провідні тканини утворюються з прокамбію в точках зростання зародка насіння та нирок відновлення. Вторинні провідні тканини, характерні для дводольних покритонасінних, породжуються камбієм.
Залежно від виконуваних функцій провідні тканини поділяються на тканини висхідного струму та тканини низхідного струму. Основним призначенням тканин висхідного струму є транспорт води та розчинених у ній мінеральних речовин від кореня до вище розташованих надземних органів. Крім того, по них переміщуються органічні речовини, що утворюються в корені та стеблі, наприклад, органічні кислоти, вуглеводи та фітогормони. Однак термін "висхідний струм" не слід сприймати однозначно як пересування знизу - вгору. Тканини висхідного струму забезпечують потік речовин у напрямку від зони всмоктування до апексу втечі. При цьому речовини, що транспортуються, використовуються як самим коренем, так і стеблом, гілками, листям, репродуктивними органами, незалежно від того, вище або нижче рівня коренів вони знаходяться. Наприклад, у картоплі вода та елементи мінерального живлення надходять по тканинах висхідного струму в столони та бульби, що утворюються у ґрунті, а також у надземні органи.
Тканини низхідного струму забезпечують відтік продуктів фотосинтезу в частини рослин і в запасні органи. У цьому просторове становище фотосинтезирующих органів немає значення. Наприклад, у пшениці органічні речовини надходять у зернівки, що розвиваються, з листя різних ярусів. Тому до назв «висхідні» і «низхідні» тканини слід відноситися не більше як до традиції, що склалася.
6.2. Провідні тканини висхідного струму
До тканин висхідного струму відносяться трахеїди та судини (трахеї), які розташовуються в деревинній (ксилемній) частині органів рослин. У цих тканинах пересування води та розчинених у ній речовин відбувається пасивно під дією кореневого тиску та випаровуванням води з поверхні рослини.
Трахеїди мають більш давнє походження. Вони зустрічаються у вищих спорових рослин, голонасінних і рідше – у покритонасінних. У покритонасінних вони типові для найдрібніших розгалужень жилок листа. Клітини трахеїд мертві. Вони мають витягнуту, часто веретеноподібну форму. Їхня довжина становить 1 – 4 мм. Однак у голонасінних, наприклад, у араукарії, вона досягає 10 мм. Стіни клітин товсті, целюлозні, часто просочуються лігніном. У клітинних оболонках є численні облямовані пори.
Судини сформувалися більш пізніх етапах еволюції. Вони характерні для покритонасінних, хоча зустрічаються і в деяких сучасних представників відділів Плауни (рід Селлагінелла), Хвощі, Папороті та Голосонасінні (рід Гнетум).
Судини складаються з подовжених мертвих клітин, розташованих одна над іншою і званих члениками судини. У торцевих стінках члеників судини є великі наскрізні отвори – перфорації, якими здійснюється далекий транспорт речовин. Перфорації виникли в ході еволюції з облямованих пір трахеїд. У складі судин вони бувають сходовими та простими. Численні сходові перфорації утворюються на торцевих стінках члеників судини за її косому закладенні. Отвори таких перфорацій мають подовжену форму, а перегородки, що їх розділяють, розташовуються паралельно один одному, нагадуючи сходи сходів. Судини зі сходовою перфорацією характерні для рослин сімейств Лютикові, Лимонникові, Березові, Пальмові, Частухові.
Прості перфорації відомі у еволюційно молодших сімейств, таких як Пасльонові, Гарбузові, Астрові, Мятлікові. Вони є одним великим отвіром в торцевій стінці членика, розташованої перпендикулярно осі судини. У ряді сімейств, наприклад, у Магнолієвих, Рожевих, Ірисових, Астрових, у судинах зустрічаються як прості, так і сходові перфорації.
Бічні стінки мають нерівномірні целюлозні потовщення, які оберігають судини від надлишкового тиску, що створюється поруч розташованими живими клітинами інших тканин. У бічних стінках можуть бути численні пори, які забезпечують вихід води межі судини.
Залежно від характеру потовщень, типів і характеру розташування пір судини поділяються на кільчасті, спіральні, біспіральні, сітчасті, сходові та точково-порові. У кільчастих та спіральних судин целюлозні потовщення розташовуються у вигляді кілець або спіралей. Через непотовщені ділянки здійснюється дифузія розчинів, що транспортуються в навколишні тканини. Діаметр цих судин порівняно невеликий. У сітчастих, сходових і точково-порових судин вся бічна стінка, за винятком місць розташування простого часу, потовщена і часто просочена лігніном. Тому радіальний транспорт речовин у них здійснюється через численні подовжені та точкові пори.
Судини мають обмежений термін. Вони можуть руйнуватися внаслідок закупорки тилами – виростами сусідніх паренхімних клітин, а також під дією доцентрових сил тиску нових клітин деревини, що утворюються камбієм. У ході еволюції судини зазнають змін. Членники судин стають коротшими і товстішими, косі поперечні перегородки змінюються прямими, а сходові перфорації - простими.
6.3. Провідні тканини низхідного струму
До тканин низхідного струму відносяться ситоподібні клітини та ситоподібні трубки з клітинами-супутницями. Ситоподібні клітини мають більш давнє походження. Вони зустрічаються у вищих спорових рослин та голонасінних. Це живі, видовжені клітини із загостреними кінцями. У зрілому стані вони містять ядра у складі протопласту. У їх бокових стінках, у місцях зіткнення суміжних клітин, є дрібні наскрізні перфорації, які зібрані групами і утворюють ситоподібні поля, якими здійснюється пересування речовин.
Ситоподібні трубки складаються з вертикального ряду подовжених клітин, розділених між собою поперечними стінками та званими ситоподібними пластинками, в яких розташовані ситоподібні поля. Якщо ситоподібна пластинка має одне ситоподібне поле, вона вважається простою, а якщо декількома – то складною. Ситоподібні поля утворюються численними наскрізними отворами – ситоподібними перфораціями невеликого діаметра. Через ці перфорації з однієї клітини до іншої проходять плазмодесми. На стінках перфорацій розміщується полісахарид калозу, що зменшує просвіт перфорацій. У міру старіння ситовидної трубки калоз повністю закупорює перфорації і трубка припиняє роботу.
При формуванні ситовидної трубки у клітинах, що їх утворюють, синтезується спеціальний флоемний білок (Ф-білок) і розвивається велика вакуоля. Вона відтісняє цитоплазму та ядро до стінки клітини. Потім мембрана вакуолі руйнується і внутрішній простір клітини заповнюється сумішшю цитоплазми та клітинного соку. Тільця Ф-білка втрачають чіткі контури, зливаються, утворюючи тяжи біля ситоподібних пластинок. Їхні фібрили проходять через перфорації з одного членика ситовидної трубки в інший. До члеників ситовидної трубки щільно прилягають одна або дві клітини-супутниці, які мають подовжену форму, тонкі стінки та живу цитоплазму з ядром та численними мітохондріями. У мітохондріях синтезується АТФ, необхідна для транспортування речовин ситоподібними трубками. У стінках клітин-супутниць є велика кількість пір з плазмадесмами, яка майже в 10 разів перевищує їх кількість в інших клітинах мезофілу листа. Поверхня протопласту цих клітин значно збільшена за рахунок численних складок, що утворюються плазмалемою.
Швидкість пересування асимілятів ситоподібними трубками значно перевищує швидкість вільної дифузії речовин і досягає 50 - 150 см/год, що вказує на активний транспорт речовин з використанням енергії АТФ.
Тривалість роботи ситоподібних трубок у багаторічних дводольних становить 1 – 2 роки. На зміну їм камбій постійно утворює нові провідні елементи. У однодольних, позбавлених камбію, ситоподібні трубки є набагато довше.
6.4. Провідні пучки
Провідні тканини розташовуються в органах рослин у вигляді поздовжніх тяжів, утворюючи пучки, що проводять. Розрізняють чотири типи провідних пучків: прості, загальні, складні та судинно-волокнисті.
Прості пучки складаються з одного типу провідних тканин. Наприклад, у крайових частинах листових пластинок багатьох рослин зустрічаються невеликі по діаметру пучки з судин і трахеїд, а в квітконосних пагонах у лілейних - лише з ситовидних трубок.
Загальні пучки утворюються трахеїдами, судинами та ситоподібними трубками. Іноді цей термін використовується для позначення пучків метамеру, які проходять у міжвузлі та є листовими слідами. До складу складних пучків входять провідні та паренхімні тканини. Найбільш досконалими, різноманітними за будовою та місцем розташування є судинно-волокнисті пучки.
Судинно-волокнисті пучки характерні для багатьох вищих спорових рослин та голонасінних. Однак вони найбільш типові для покритонасінних. У таких пучках виділяються функціонально різні частини – флоема та ксилема. Флоема забезпечує відтік асимілятів з листа та пересування їх у місця використання чи запасання. По ксилемі вода та розчинені в ній речовини пересуваються з кореневої системи в лист та інші органи. Об'єм ксилемної частини в кілька разів перевищує обсяг флоемної, оскільки обсяг води, що надходить в рослину, перевищує обсяг утворюваних асимілятів, так як значна частина води випаровується рослиною.
Різноманітність судинно-волокнистих пучків визначається їх походженням, гістологічним складом та місцезнаходженням у рослині. Якщо пучки утворюються з прокамбію і завершують свій розвиток у міру використання запасу клітин освітньої тканини, як у однодольних, вони називаються закритими для зростання. На відміну від них, у дводольних відкриті пучки не обмежені у зростанні, оскільки вони формуються камбієм і збільшуються в діаметрі протягом усього життя рослини. До складу судинно-волокнистих пучків крім провідних можуть входити основні та механічні тканини. Наприклад, у дводольних флоема утворюється ситовидними трубками (провідна тканина висхідного струму), луб'яною паренхімою (основна тканина) і луб'яними волокнами (механічна тканина). До складу ксилеми входять судини та трахеїди (що проводить тканину низхідного струму), деревна паренхіма (основна тканина) та деревні волокна (механічна тканина). Гістологічний склад ксілеми та флоеми генетично детермінований і може бути використаний у систематиці рослин для діагностики різних таксонів. Крім того, ступінь розвитку складових частин пучків може змінюватись під впливом умов зростання рослин.
Відомо кілька видів судинно-волокнистих пучків.
Закриті колатеральні провідні пучки характерні для листя та стебел однодольних покритонасінних. Вони відсутні камбій. Флоема і ксилема розташовуються пліч-о-пліч. Їх характерні деякі конструктивні особливості. Так, у пшениці, що відрізняється С 3 шляхом фотосинтезу, пучки утворюються з прокамбію і мають первинну флоему і первинну ксилему. У флоемі виділяють більш ранню протофлоему і пізнішу за часом утворення, але більш великоклітинну метафлоему. У флоемній частині відсутні луб'яна паренхіма та луб'яні волокна. У ксілемі спочатку утворюються дрібніші судини протоксилеми, розташовані в одну лінію перпендикулярно до внутрішньої межі флоеми. Метаксилема представлена двома великими судинами, розташованими поруч із метафлоемою перпендикулярно ланцюжку судин протоксилеми. І тут судини розташовуються Т-образно. Відомо також V-, Y- та È-подібне розташування судин. Між судинами метаксилеми в 1 - 2 ряди розташована дрібноклітинна склеренхіма з потовщеними стінками, які в міру розвитку стебла просочуються лігніном. Ця склеренхіма відокремлює зону ксілеми від флоеми. По обидва боки від судин протоксилеми розташовуються клітини деревинної паренхіми, які, ймовірно, виконують трансфузійну роль, оскільки при переході пучка з міжвузля в листову подушку стеблового вузла вони беруть участь в утворенні клітин. Навколо пучка стебла пшениці, що проводить, розташовується склеренхімна обкладка, краще розвинена з боку протоксилеми і протофлоеми, біля бічних сторін пучка клітини обкладки розташовуються в один ряд.
У рослин з 4-типом фотосинтезу (кукурудза, просо та ін) в листі навколо закритих провідних пучків розташовується обкладка з великих клітин хлоренхіми.
Відкриті колатеральні пучки характерні для дводольних стебел. Наявність шару камбію між флоемою та ксилемою, а також відсутність склеренхімної обкладки навколо пучків забезпечує їх тривале зростання у товщину. У ксилемній та флоемній частинах таких пучків є клітини основної та механічної тканин.
Відкриті колатеральні пучки можуть бути утворені двома шляхами. По-перше, це пучки, які первинно утворюються прокамбієм. Потім у них із клітин основної паренхіми розвивається камбій, що виробляє вторинні елементи флоеми та ксилеми. В результаті пучки поєднують гістологічні елементи первинного та вторинного походження. Такі пучки характерні для багатьох трав'янистих квіткових рослин класу дводольних, що мають пучковий тип будови стебла (бобові, рожеві та ін.).
По-друге, відкриті колатеральні пучки можуть бути утворені лише камбієм і складатися з ксилеми та флоеми вторинного походження. Вони типові для трав'янистих дводольних з перехідним типом анатомічної будови стебла (астрові та ін), а також для коренеплодів типу буряків.
У стеблах рослин ряду сімейств (Гарбузові, Пасльонові, Дзвіночкові та ін) зустрічаються відкриті біколатеральні пучки, де ксилема з двох сторін оточена флоемою. При цьому зовнішня ділянка флоеми, звернена до поверхні стебла, розвинена краще внутрішнього, а смужка камбію, як правило, розташовується між ксилемою і зовнішньою ділянкою флоеми.
Концентричні пучки бувають двох типів. У амфікрибральних пучках, характерних для кореневищ папороті, флоема оточує ксилему, в амфівазальних – ксилема кільцем розташована навколо флоеми (кореневища ірису, конвалії та ін.). Рідше концентричні пучки зустрічаються у дводольних (кліщевина).
Закриті радіальні провідні пучки утворюються в ділянках коренів, що мають первинну анатомічну будову. Радіальний пучок входить до складу центрального циліндра та проходить через середину кореня. Його ксилема має вигляд багатопроменевої зірки. Між променями ксілеми розташовуються клітини флоеми. Число променів ксілеми значною мірою залежить від генетичної природи рослин. Наприклад, у моркви, буряків, капусти та інших дводольних ксилема радіального пучка має лише два промені. У яблуні та груші їх може бути 3 – 5, у гарбуза та бобів – ксилема чотирипроменева, а у однодольних – багатопроменева. Радіальне розташування променів ксілеми має пристосувальне значення. Воно скорочує шлях води від поверхні, що всмоктує, кореня до судин центрального циліндра.
У багаторічних деревних рослин і деяких трав'янистих однорічників, наприклад, у льону, провідні тканини розташовуються в стеблі, не утворюючи чітко виражених провідних пучків. Тоді говорять про непучковий тип будови стебла.
6.5. Тканини, що регулюють радіальний транспорт речовин
До специфічних тканин, що регулюють радіальний транспорт речовин відносяться екзодерми та ендодерма.
Екзодерма є зовнішнім шаром первинної кори кореня. Вона утворюється безпосередньо під первинною покривною тканиною епіблемою в зоні кореневих волосків і складається з одного або декількох шарів щільно зімкнутих клітин з потовщеними целюлозними оболонками. В екзодермі вода, що надійшла в корінь кореневими волосками, відчуває опір в'язкої цитоплазми і переміщається в целюлозні оболонки клітин екзодерми, а потім виходить з них в міжклітини середнього шару первинної кори, або мезодерми. Це забезпечує ефективне надходження води у глибші шари кореня.
У зоні проведення корені однодольних, де клітини епіблеми відмирають і злущуються, екзодерма виявляється на поверхні кореня. Її клітинні стінки просочуються суберином і перешкоджають надходженню води із ґрунту в корінь. У дводольних екзодерми у складі первинної кори злущується при линянні кореня і заміщається перидермою.
Ендодерма, або внутрішній шар первинної кори кореня, розташований навколо центрального циліндра. Вона утворюється одним шаром щільно зімкнутих клітин неоднакової будови. Одні з них, названі пропускними, мають тонкі оболонки і легко проникні для води. За ними вода з первинної кори надходить у радіальний провідний пучок кореня. Інші клітини мають специфічні целюлозні потовщення радіальних та внутрішніх тангентальних стінок. Ці потовщення, просочені суберином, називаються поясками Каспарі. Вони непроникні для води. Тому вода надходить у центральний циліндр лише через пропускні клітини. А оскільки поглинаюча поверхня кореня значно перевищує сумарну площу перерізу пропускних клітин ендодерми, то при цьому виникає кореневий тиск, який є одним із механізмів надходження води в стебло, лист та репродуктивні органи.
Ендодерма входить також до складу кори молодого стебла. У деяких трав'янистих покритонасінних вона, як і в корені, може мати пояски Каспарі. Крім того, в молодих стеблах ендодерма може бути представлена крохмалоносною піхвою. Таким чином, ендодерма може регулювати транспорт води у рослині та запасати поживні речовини.
6.6. Поняття про стелу та її еволюцію
Виникненню, розвитку в онтогенезі та еволюційним структурним перетворенням провідної системи приділяється велика увага, оскільки вона забезпечує взаємозв'язок органів рослин та з нею пов'язана еволюція великих таксонів.
На пропозицію французьких ботаніків Ф. Ван Тигема і А. Дуліо (1886) сукупність первинних провідних тканин разом із іншими тканинами і перициклом, що прилягають до кори, була названа стелою. До складу стели може також входити серцевина і утворена її місці порожнину, як, наприклад, у мятликових. Поняття "стела" відповідає поняттю "центральний циліндр". Стела кореня та стебла функціонально єдина. Вивчення стели у представників різних відділів вищих рослин призвело до формування стелярної теорії.
Розрізняють два основні типи стели: протостелу та еустелу. Найбільш давньою є протостела. Її провідні тканини розташовуються в середині осьових органів, причому в центрі знаходиться ксилема, оточена суцільним шаром флоеми. Серцевина чи порожнина у стеблі відсутні.
Існує кілька еволюційно пов'язаних між собою видів протостели: гаплостела, актиностела та плектостела.
Вихідним, примітивним виглядом є гаплостели. У неї ксилема має округлу форму поперечного перерізу і оточена безперервним рівним шаром флоеми. Навколо провідних тканин одним - двома шарами розташовується перицикл [К. Есау, 1969]. Гаплостела була відома у копалин рініофітів і збереглася у деяких псилотофітів (тмезіптер).
Більш розвиненим видом протостели є актиностела, в якій ксилема на поперечному перерізі набуває форми багатопроменевої зірки. Вона виявлена у викопного астероксилону та деяких примітивних плауноподібних.
Подальше роз'єднання ксилеми на окремі ділянки, розташовані радіально або паралельно один до одного, призвело до утворення плектостели, характерної для плауноподібних стебел. У актиностели та плектостели флоема, як і раніше, оточує ксилему з усіх боків.
У ході еволюції з протостели виникла сифоностела, відмінною рисою якої є трубчаста будова. У центрі такої стели розташовується серцевина чи порожнина. У провідній частині сифоностели з'являються листові щілини, завдяки яким виникає безперервний зв'язок серцевини з корою. Залежно від способу взаємного розташування ксилеми і флоеми сифоностела буває ектофлою та амфіфлою. У першому випадку флоема з однієї, зовнішньої сторони оточує ксилему. У другому – флоема оточує ксилему із двох сторін, із зовнішньої та внутрішньої.
При поділі амфіфлойної сифоностели на мережу або ряди поздовжніх тяжів виникає розсічена стела, або диктіостела, характерна для багатьох папоротеподібних. Її провідна частина представлена численними концентричними пучками, що проводять.
У хвощів з ектофлойної сифоностели виникла артростела, яка має членисту будову. Вона відрізняється наявністю однієї великої центральної порожнини та відокремлених провідних пучків з протоксилемними порожнинами (каринальними каналами).
У квіткових рослин на основі ектофлойної сифоностели утворилася еустела, характерна для дводольних, і атактостела, типова для однодольних. В еустелі провідна частина складається з відокремлених колатеральних пучків, що мають кругове розташування. У центрі стели в стеблі розташовується серцевина, яка за допомогою серцевих променів з'єднується з корою. В атактостелі провідні пучки мають розсіяне розташування, між ними знаходяться паренхімні клітини центрального циліндра. Таке розташування пучків приховує трубчасту конструкцію сифоностели.
Виникнення різних варіантів сифоностели є важливим пристосуванням вищих рослин до збільшення діаметра осьових органів – кореня та стебла.
25. 8.1. Видільна система рослин та її значення
Життя рослин є генетично детермінованою сукупністю біохімічних реакцій, швидкість і інтенсивність яких значною мірою модифікується умовами середовища проростання. У цих реакціях утворюється велика різноманітність побічних продуктів, що не використовуються рослиною для побудови тіла або регулювання обміну речовинами, енергією та інформацією з навколишнім середовищем. Такі продукти можуть видалятися з рослини різними способами: при відмиранні та відділенні гілок та ділянок кореневищ, при опаданні листя та злущуванні зовнішніх шарів кірки, в результаті діяльності спеціалізованих структур зовнішньої та внутрішньої секреції. У сукупності ці пристрої утворюють видільну систему рослин.
На відміну від тварин система виділення у рослин не спрямована на видалення сполук азоту, який може реутилізуватися в процесі життєдіяльності.
Видільна система рослин багатофункціональна. У її структурах здійснюються: синтез, накопичення, проведення та виділення продуктів метаболізму. Наприклад, у секреторних клітинах смоляних ходів у листі хвойних порід утворюється смола, що виділяється через смоляні ходи. У нектарниках квіток липи утворюється та виділяється солодкий сік нектар. У спеціальних вмістилищах у плодовій оболонці у цитрусових накопичуються ефірні масла.
Утворення та виділення побічних продуктів метаболізму має різноманітне пристосувальне значення:
Залучення комах-запилювачів. У квітках яблуні, огірка та інших ентомофільних перехреснозапилювачів утворюється нектар, що приваблює бджіл, а смердючі виділення квітки раффлезії приваблюють мух;
Відлякування травоїдних тварин (кмин, кропива та ін);
Захист від бактерій та грибів, що руйнують деревину (сосна, ялина та ін.);
Виділення в атмосферу летких сполук, що сприяє очищенню повітря від хвороботворних бактерій;
Позаклітинне перетравлення видобутку у комахоїдних рослин рахунок виділення протеолітичних ферментів (росянка, альдрованда та інших.);
Мінералізація органічних залишків у ґрунті завдяки виділенню корінням спеціальних ґрунтових ферментів;
Регулювання водного режимуза допомогою водяних продихів - гідатод, розташованих по краю листової пластинки (суниця, капуста, товстянка та ін);
Регулює випаровування води в результаті виділення летких ефірних сполук, які зменшують прозорість і теплопровідність повітря біля поверхні листа (хвойні породи);
Регулювання сольового режиму клітин (мар, лобода та ін);
Зміна хімічних та фізичних властивостейґрунти, а також регулювання видового складу ґрунтової мікрофлори під впливом кореневих виділень;
Регулювання взаємодії рослин у фітоценозі за допомогою кореневих, стеблових та листових виділень, що називається алелопатією (цибуля, часник та ін.).
Виділяються рослинами речовини дуже різноманітні. Їхня природа залежить від генотипу рослин.
Багато видів виділяють воду (суниця, капуста), солі (мар, лобода), моносахариди та органічні кислоти (кульбаба, цикорій), нектар (липа, гречка), амінокислоти та білки (тополі, верби), ефірні олії (м'ята, троянда) , бальзам (ялиця), смоли (сосна, ялина), каучук (гевея, кок-сагиз), слизу (клітини кореневого чохлика, набухають насіння різних рослин), травні соки (росянка, жирянка), отруйні рідини (кропива, борщівка) інші сполуки.
26. 1.1. Полярність
Полярність – це біохімічних, функціональних і структурних відмінностей у діаметрально протилежних частинах органів цілісного рослинного організму. Полярність впливає інтенсивність біохімічних процесів у клітині і функціональну активність органоїдів, визначає конструкцію анатомічних структур. На рівні цілісного рослинного організму з полярністю пов'язана спрямованість зростання та розвитку за умов дії гравітаційних сил.
Явище полярності спостерігається різних рівнях організації рослин. На молекулярному рівні вона проявляється у будові молекул органічних речовин, насамперед – нуклеїнових кислот та білків. Так, полярність ланцюжків ДНК визначається спеціальним порядком з'єднання її нуклеотидів. Полярність поліпептидів пов'язана з наявністю у складі амінокислот аміногрупи –NH 2 та карбоксильної групи –COOH. Полярність молекул хлорофілу обумовлена наявністю в ній тетрапіррольного порфіринового ядра та залишків спиртів – метанолу та фітолу.
Полярністю можуть мати клітини та їх органоїди. Наприклад, на регенераційному полюсі комплексу Гольджі утворюються нові диктіосоми, а на секреторному – бульбашки, які пов'язані з виведенням із клітини продуктів метаболізму.
Полярність клітин виникає у процесі їхнього онтогенезу. Поляризація клітин відбувається внаслідок виникнення градієнтів pH, електричних зарядівта осмотичного потенціалу, концентрації О 2 та СО 2 , катіонів кальцію, фізіологічно активних речовин та елементів мінерального харчування. Вона може виникнути під впливом механічного тиску, сил поверхневого натягу, впливу сусідніх клітин.
Полярність клітин буває симетричною та несиметричною. Симетрична полярність є неодмінною умовою поділу вихідних та утворення рівноцінних дочірніх клітин. Зокрема, мікротрубочки цитоскелета переміщуються до екваторіальної площини та беруть участь в утворенні фрагмопласту; Хромосоми діляться і їх хроматиди за допомогою ниток, що тягнуть, ахроматинового веретена розходяться до протилежних полюсів клітини. Тут же починають концентруватися рибосоми та мітохондрії. Комплекси Гольджі переміщуються до центру та специфічно орієнтуються у просторі. Їх секреторні полюси спрямовані в бік екватора клітини, що ділиться, що забезпечує активну участь цього органоїду в утворенні серединної пластинки і первинної клітинної оболонки.
Несиметрична полярність й у спеціалізованих клітин. Так, у зрілій яйцеклітині ядро зміщене до полюса, орієнтованого до халази сім'язачатка, а велика вакуоль розташовується біля мікропілярного полюса.
Ще сильніша несиметрична полярність виражена в заплідненій яйцеклітині - зиготі. У ній ядро серед канальців ендоплазматичної мережі розташовується з халазальної боку клітини. Тут більш щільно розміщуються елементи цитоскелета, підвищується оптична щільність цитоплазми, відзначається підвищений вміст ферментів, фітогормонів та інших фізіологічно активних речовин. Функціонально активніша її частина стає апікальною, а протилежна – базальною.
З апікальної клітини при розвитку зародка насіння утворюються зародкова стеблинка з конусом наростання і примордіальним листям. З базальної клітини формується підвісок, пізніше - зпродишевий корінець.
Завдяки полярності клітин зародка, що розвивається, зріле насіння також виявляється поляризованим. Наприклад, у насінні пшениці з одного боку розташований ендосперм, а з іншого – зародок, у якому апікальна меристема стеблинки та кінчика корінця знаходяться на різних полюсах.
На відміну від клітин, полярність органів рослин має більш універсальний характер. Вона проявляється у структурі втечі та кореневої системи, і навіть їх складових частин. Наприклад, базальна та апікальна частини втечі розрізняються морфологічно, анатомічно, гістологічно, біохімічно та функціонально. У багатьох квіткових рослин верхівці пагона знаходиться верхівкова нирка, у якій розташований найважливіший морфогенетичний центр рослини – конус наростання. Завдяки його діяльності утворюються зачатки листя, бічних пазушних бруньок, вузлів та міжвузлів. Диференціювання клітин первинної меристеми призводить до виникнення первинних покривних тканин, гістологічних елементів первинної кори та центрального циліндра.
Верхівка втечі є потужним центром, що атрагує. Сюди прямує основний потік води та розчинених у ній елементів мінерального живлення та органічних речовин, що синтезуються в корені. Переважне надходження цитокініна до верхівкової нирки призводить до апікального домінування. Клітини апекса верхівкової бруньки активно діляться, забезпечуючи зростання стебла в довжину та утворення нового листя та пазушних бруньок на головній осі. У цьому спостерігається корелятивне гальмування розвитку пазушних нирок.
Гарним прикладомМорфологічною полярністю втечі є будова стебла пшениці. У міру переміщення знизу - вгору, від базальної частини втечі до апікальної, міжвузля стають довшими, їх товщина в середній частині спочатку збільшується, а потім поступово зменшується. У листя верхнього ярусу відношення ширини листової пластинки до її довжини значно більше, ніж у листя нижнього ярусу. Змінюється також положення листя у просторі. У пшениці листя нижнього ярусу поникають вниз, листя середнього ярусу розташовується майже плагіотропно, тобто. паралельно поверхні землі, а прапорне (верхнє) листя прагне до ортотропного - майже вертикального, положення.
Полярність втечі добре простежується на анатомічному рівні. У видів пшениці підколосове міжвузля в порівнянні з нижче розташованими міжвузлями відрізняється меншим діаметром, меншою кількістю пучків, що проводять, краще розвиненою асиміляційною паренхімою.
У дводольних рослин анатомічна полярність втечі посилюється завдяки виникненню вторинних бічних освітніх тканин – камбію та феллогену. Феллоген породжує вторинну покривну тканину фелему, яка, просочуючись суберином, перетворюється на пробку. У деревних порід діяльність феллогену призводить до утворення третинного покривного комплексу – кірки. Камбій забезпечує перехід до вторинної анатомічної будови стебла нижче рівня розташування верхівкової бруньки.
Для коренів, як і інших органів рослин, також характерна структурна і функціональна полярність. У зв'язку з позитивним геотропізмом базальна частина кореня знаходиться біля поверхні ґрунту. Вона безпосередньо пов'язана з кореневою шийкою – місцем переходу кореня до стебла. Апікальна частина кореня зазвичай заглиблена у ґрунт. У ній проходять взаємопов'язані процеси зростання та розвитку. Розподіл клітин меристеми апекса забезпечує лінійний приріст кореня. А в результаті диференціації клітин утворюються якісно нові структури. На клітинах епіблем з'являються кореневі волоски. З клітин периблеми формуються елементи первинної кори, та якщо з плероми – перицикл і провідні елементи центрального циліндра. Така конструкція молодих ділянок кореня забезпечує активне всмоктування води та мінеральних речовин, а також подачу їх у вище розташовані ділянки кореня. Функції базальної частини кореня дещо відрізняються. Так, у багаторічних дводольних базальна частина кореня виконує транспортну, опорну та запасну функцію. Виконання цих функцій відповідає анатомічна будова. Тут краще розвинені провідні тканини, кора утворюється завдяки діяльності камбію, покривна тканина представлена пробкою. Полярність структури кореня забезпечує різноманітність його функцій.
Полярність характерна й у репродуктивних органів рослин. Так, квітка, будучи видозміненою укороченою втечею, зберігає ознаки полярності втечі. На ньому в закономірній послідовності розташовуються частини квітки: філіжанка, віночок, андроцей і гинецей. Таке розташування сприяє кращому вловлюванню пилку приймочком маточка, а також забезпечує захист генеративних частин квітки вегетативними. Дуже показовою є полярність суцвіть. У невизначених суцвіттях першими закладаються квіти базальної частини. Вони досягають великих розміріві утворюють плоди з добре розвиненим насінням. Зокрема, більше насіння у соняшника утворюється в периферичній частині кошика, а в складному колосі пшениці. найкраще насінняутворюються у перших квітках колосків. Квітки апікальної частини суцвіть формуються пізніше. Одержувані з них плоди і насіння бувають дрібнішими, а їх посівні якості – нижче.
Таким чином, полярність – це важлива конструктивна, функціональна та біохімічна особливість рослин, що має адаптивне значення та яку необхідно враховувати в агрономічній практиці.
1.2. Симетрія
Навколишній світ характеризується цілісністю і гармонійною впорядкованістю. У найближчому космічному просторі гармонійні положення, маса, форма та траєкторія руху об'єктів Сонячної системи. На Землі гармонійні сезонні та добові зміни найважливіших фізичних параметрів умов існування живих організмів, котрим також характерне гармонійне узгодження будови та функцій. Мудреці школи Піфогора вважали, що гармонія – це «спосіб узгодження багатьох частин, з допомогою якого об'єднуються в ціле». Відображенням гармонії у природі є симетрія. За визначенням Ю.А. Урманцева симетрія – це категорія, що означає збереження ознак об'єктів щодо їх змін. В утилітарному плані симетрія є однотипністю будови і взаємного розташування однотипних складових частин єдиного цілого. Симетрія притаманна як мінералів, так об'єктів живої природи. Однак форми симетрії та ступінь їхнього прояву у різних об'єктів значно різняться.
Істотною особливістю симетрії різних за походженням об'єктів є співвідносний характер. Порівняння структур проводиться у кількох характерних точках. Точки фігури, які дають ту саму картину при розгляді фігури з різних сторін, називаються рівними. Це можуть бути рівновіддалені точки на прямій, точки перетину сторін рівнобедреного трикутника і квадрата, граней багатогранника, точки на колі або поверхні кулі і т.д.
Якщо фігурі для деякої точки Х немає іншої рівної точки, вона називається особливої. Фігури, що мають одну особливу та кілька рівних точок отримали назву розетки. Фігури, що не містять рівних точок, вважаються асиметричними.
Через спеціальні точки проходять осі і поверхні симетрії. У рівнобедреному трикутнику може бути проведено три, у квадраті – чотири, у рівносторонньому п'ятикутнику – п'ять площин симетрії. Відповідно, на прямій утворюється два промені симетрії, для трикутника характерна трипроменева симетрія, для квадрата – чотирипроменева, для кола – багатопроменева.
Рухи фігури, у яких кожна точка заміщається рівної їй точкою, а кожна особлива точка залишається дома, називаються перетвореннями симетрії, а фігури, котрим допустиме перетворення симетрії, вважаються симетричними.
Найбільш поширеними формами перетворення симетрії є:
1. Відображення – переміщення кожної точки, що знаходиться на деякій відстані від нерухомої площини, по прямій, перпендикулярній цій площині, на таку ж відстань по інший бік від неї (наприклад, дзеркальна симетрія квітки зигоморфного гороху);
2. Поворот - переміщення всіх точок на деякий кут навколо нерухомої осі (наприклад, багатопроменева симетрія актиноморфного квітки вишні);
3. П а р а л е л ь н ий пе ренос, наприклад, розташування метамерів на втечі.
Форми перетворення симетрії є тотожними і зводяться одна до одної. Так, у разі відображення нерухомої залишається площина; при повороті – пряма (вісь); а при паралельному перенесенні жодна точка залишається на місці.
Симетричні фігури з кількома точками можуть мати різне число особливих точок. Так, розетка має одну особливу точку. Основною формою перетворення симетрії нею є поворот. Для фігур, які мають особливих точок, характерний паралельний перенесення, чи зсув. Такі постаті умовно називають нескінченними.
2.2.1. Особливості вияву симетрії у рослин. Симетрія рослин відрізняється від симетрії кристалів рядом особливостей.
1) Симетрія рослин визначається не тільки симетрією молекул, що утворюють їх клітини, а й симетрією умов довкілля, у яких відбувається розвиток рослин.
Приватними випадками симетрії довкілля вважатимуться однорідність складання корнеобитаемого шару грунту, рівномірність розподілу у грунті води та елементів мінерального харчування, рівна віддаленість рослин, що ростуть поруч. Ці обставини повинні враховуватися під час вирощування культурних рослин.
2) У рослин, як та інших живих організмів, немає абсолютної тотожності елементів складових частин. Це насамперед залежить від того, що умови формування цих елементів є абсолютно тотожними. З одного боку, це пов'язано з різночасністю вичленування конусом наростання різних метамерів, що включають зачатки листя, нирок, вузлів та міжвузлів. Це саме стосується й генеративних органів. Так, крайові трубчасті квітки в кошику соняшнику завжди більші за квітки центральної частини суцвіття. З іншого боку, протягом вегетації спостерігається як закономірна, і випадкова зміна умов розвитку. Наприклад, статистично закономірно протягом вегетаційного періоду змінюється температура повітря та ґрунту, освітленість та спектральний склад світла. Випадковими можуть бути локальні зміни запасу поживних речовин у ґрунті, дії хвороб та шкідників.
Відсутність абсолютної тотожності складових частин має значення у житті рослин. Виникає гетерогенність одна із механізмів надійності онтогенезу.
3) Для рослин характерна субстанційна, просторова та тимчасова симетрія.
А. С у б с т а н ц і о н а л ь н а я з і метра полягає в точному повторенні форми та лінійних параметрів структур, а також у точній зміні цих параметрів. Вона характерна для кристалів неорганічних речовин, молекул органічних сполук, органоїдів клітини, анатомічних структур та органів рослин. Високим рівнемСубстанційною симетрією відрізняються молекули ДНК. При вологості препаратів близької до фізіологічної молекула ДНК знаходиться в В-формі і характеризується чітким повторенням параметрів її складових елементів. Кожен виток спіралі молекули ДНК містить 10 нуклеотидів; проекція витка на вісь молекули становить 34,6 Å (1 Å = 1 · 10 -10 м); проекційна відстань між сусідніми нуклеотидами дорівнює 3,4 Å, а лінійна – 7 Å; діаметр молекули при орієнтуванні атомами фосфору близький до 20 Å; діаметр великої борозни приблизно дорівнює 17 Å, а малої - 11 Å.
Субстанційна симетрія рослин характеризується як точним повторенням параметрів структур, а й закономірним їх зміною. Наприклад, у ялини зменшення діаметра ствола, що припадає на 1 м довжини, при переміщенні від його базальної частини до верхівки є відносно постійним. У канни фіолетової (Canna violacea) квітка традиційно вважається асиметричною. У нього чашолистки, пелюстки та стамінодії мають різні розміри. Однак зміна лінійних розмірів цих членів квітки залишається відносно постійною, що є ознакою симетричності. Тільки тут замість дзеркальної розвиваються інші форми симетрії.
Б. Просторанність в я метрі полягає в закономірно повторюваному просторовому розміщенні однотипних складових частин у рослин. Просторова симетрія поширена у світі рослин. Вона характерна для розташування нирок на пагоні, квіток у суцвітті, членів квітки на квітколожі, пилку в пильовику, лусок у шишках голонасінних та багатьох інших випадків. Приватним випадком просторової симетрії є поздовжня, радіальна та змішана симетрія стебла.
Поздовжня симетрія виникає при паралельному перенесенні, тобто. просторовому повторенні метамерів у структурі втечі Вона визначається конструктивним подібністю складових частин втечі – метамерів, і навіть відповідністю довжини междоузлий правилу «золотого перерізу».
Більш складним випадком просторової симетрії є розташування листя на втечі, яке може бути мутовчастим, супротивним та черговим (спіральним).
Радіальна симетрія виникає при комбінуванні осі структури з площинами симетрії, що проходять через неї. Радіальна симетрія поширена у природі. Зокрема вона характерна для багатьох видів діатомових водоростей, поперечного зрізу стебла вищих спорових рослин, голонасінних та покритонасінних.
Радіальна симетрія втечі є багатоелементною. Вона характеризує місцезнаходження складових частин і може бути виражена різними мірними показниками: віддаленістю провідних пучків та інших анатомічних структур від особливої точки, через яку проходять одна або кілька площин симетрії, ритмічністю чергування анатомічних структур, кутом дивергенції, який показує зміщення осі однієї структури по відношенню до осі інший. Наприклад, у квітці вишні просторово симетрично розташовуються п'ять пелюсток віночка. Місце прикріплення кожного з них до кольороложа знаходиться на однаковій відстані від центру квітки, а кут дивергенції дорівнюватиме 72º (360º: 5 = 72º). У квітки тюльпана кут дивергенції кожного з шести пелюсток становить 60 º (360 º: 6 = 60 º).
С. В часи м е т р і я рослин виражається в ритмічному повторенні в часі процесів морфогенезу та інших фізіологічних функцій. Наприклад, вичленування конусом наростання листових примордій здійснюється через більш менш рівні проміжки часу, іменовані пластохроном. Дуже ритмічно повторюються сезонні зміни процесів життєдіяльності у багаторічних полікарпічних рослин. Тимчасова симетрія відображає адаптацію рослин до добових та сезонних змін умов довкілля.
4) Симетрія рослин динамічно розвивається в онтогенезі і досягає максимального вираження у пору статевого розмноження. У рослин цьому відношенні показовий приклад формування радіальної симетрії втечі. Спочатку клітини конуса наростання більш менш однорідні, не диференційовані. Тому тут морфологічно важко виділити клітину чи групу клітин, якими можна було б провести площину симетрії.
Пізніше клітини туніки утворюють протодерму, з якої формується епідерміс. З основної меристеми розвиваються асимілюючі та запасні тканини, а також первинні механічні тканини та серцевина. У периферичній зоні формуються тяжі вузьких і довгих клітин прокамбію, з яких розвиватимуться провідні тканини. При суцільному закладенні прокамбію з нього утворюються суцільні шари флоеми та ксилеми. Якщо ж прокамбій закладається у вигляді тяжів, то з нього формуються відокремлені пучки, що проводять. У деревних рослин багаторічна, функціональна активність камбію, що сезонно змінюється, приведе до утворення вторинної кори і річних кілець деревини, що посилить радіальну симетричність стебла.
5) Симетрія рослин змінюється під час еволюції. Еволюційні перетворення симетрії мають величезне значенняу розвитку органічного світу. Так, виникнення двосторонньої симетрії стало великим морфофізіологічним пристосуванням (ароморфозом), що значно підняв рівень організації тварин. У царстві рослин зміна числа основних видів симетрії та їх похідних пов'язана з появою багатоклітинності та виходом рослин на сушу. У одноклітинних водоростей, зокрема діатомових, широко поширена радіальна симетрія. У багатоклітинних з'являються різні формипаралельного перенесення, і навіть змішані форми симетрії. Наприклад, у харових водоростей радіальна симетрія поперечного зрізу талому поєднується з наявністю осі перенесення метамерів.
Велике значення у формуванні симетрії вищих рослин мала еволюція провідної системи. Радіально симетрична протостела стебла примітивних форм була замінена комплексом двосторонньосиметричних (моносиметричних) колатеральних пучків, що формують еустелу і забезпечують радіальну симетрію стебла у покритонасінних.
Генеральною тенденцією еволюції симетрії у живих організмів, включаючи рослини, є зменшення рівня симетричності. Це з зменшенням числа основних видів симетрії та його похідних. Так, для еволюційно більш ранніх квіткових рослин (сім. Магнолієві, сем. лютикові та ін) характерні багаточлені, вільні, спірально розташовані частини квітки. Квітка у своїй виявляється актиноморфним, тобто. полісиметричним. Формули таких квіток прагнуть виразу: Å Са ¥ Со ¥ А ¥ G ¥ . У результаті еволюції спостерігається редукція числа членів квітки, і навіть їх зрощення, що незмінно призводить до зменшення кількості площин симетрії.
В еволюційно молодих сімействах, наприклад у ясноткових або мятликових, квітки стають зигоморфними (моносиметричними). Дисиметризація квіток має явно пристосувальне значення, пов'язане з удосконаленням способів запилення. Цьому часто сприяла пов'язана еволюція квітки та запилювачів – комах та птахів.
Велике значення набула спеціалізація симетрії квіток у суцвітті. Так, у кошику соняшнику безплідні крайові зигоморфні квітки мають хибну язичок. жовтого кольору, утворений трьома пелюстки, що зрослися. Плодні квітки центральної частини суцвіття актиноморфні, вони утворені п'ятьма невеликими рівновеликими пелюстками, що зрослися в трубочку. Іншим прикладом можуть бути представники сімейства Селерні. У них у складній парасольці крайові квітки слабо зігоморфні, тоді як інші квітки залишаються типово актиноморфними.
Еволюція структури рослин та його симетрії не прямолінійна. Переважна за роль дисиметризація змінюється та доповнюється на окремих етапах еволюції симетризацією.
Таким чином, симетрія рослин та їх складових частин дуже багатогранна. Вона пов'язана з симетрією елементів, що її утворюють на молекулярному, клітинному, гістолого-анатомічному і морфологічному рівні. Симетрія динамічно розвивається в ході онтогенезу та філогенезу та забезпечує зв'язок рослин з навколишнім середовищем.
1.3. Метамірність
Важливим морфофізіологічним пристосуванням рослин є метамерність, яка є наявність повторюваних елементарних кінцевих структур, або метамерів, в системі цілісного організму. Метамерна конструкція забезпечує багаторазове повторення складових частин втечі і, отже, одна із механізмів надійності онтогенезу. Метамерна конструкція й у різних систематичних груп рослин. Вона відома у харових водоростей, хвощів та інших вищих спорових рослин, у голонасінних, наземних та водних покритонасінних. Метамерними бувають функціонально різні частини рослин – вегетативна та генеративна. За участю метамерно влаштованих органів рослин формується метамерна система. Окремим випадком утворення метамерних систем є розгалуження.
До складу метамеру вегетативної зони втечі покритонасінних входять лист, вузол, міжвузля та бічна пазушна нирка, яка розташовується біля основи міжвузля з боку, протилежної до місця прикріплення листа. Цю нирку прикриває лист попереднього метамеру. Метамери генеративної зони дуже різноманітні. Наприклад, у пшениці метамери складного колоса складаються з членика колосового стрижня та прикріпленого до нього колоска. Іноді рослин виділяється перехідна зона втечі. У деяких видів та сортів пшениці вона може бути представлена лусками недорозвинених колосків.
Метамерність рослин - це морфологічне вираження специфіки їх росту та формоутворення, які протікають ритмічно, у вигляді повторюваних супідрядних циклів, локалізованих в осередках меристеми. Ритмічність утворення метамерів перебуває у нерозривному зв'язку з періодичністю ростових процесів, притаманних рослин. Освіта метамерів є первинним морфогенетичним процесом у розвитку рослин. Воно становить основу ускладнення організації в онтогенезі і відбиває процес полімеризації, що є одним з механізмів еволюції вищих рослин.
Утворення та розвиток метамерів забезпечується насамперед функцією апікальної та інтеркалярної меристем.
У вегетативної нирці, а також у зародку насіння, що проростає, в результаті мітотичного поділу відбувається збільшення обсягу конуса наростання зародкового стеблинки. Подальше активне розподіл клітин периферичної зони конуса наростання призводить до утворення зачатка листа – листового примордя та інсерційного диска. У цей час меристематична активність туніки та центральної меристематичної зони дещо знижується, але активно діляться клітини інсерційного диска. Верхня частина диска є місцем кріплення листового примордя і при розростанні завтовшки з нього утворюється вузол. З нижньої частини інсерційного диска розвивається міжвузля. Тут же формується зародковий горбок бічної пазушної нирки з боку, протилежної до середньої жилки примордіального листа. У сукупності зачаток листа, інсерційний диск і зачаток нирки є метам зачатків.
У міру завершення утворення зародкового метамеру знову підвищується активність поділу клітин туніки та центральної меристематичної зони. Об'єм гладкої частини конуса наростання знову збільшується, досягаючи максимуму перед закладенням чергового листового примордя. Таким чином, на апікальній частині попереднього зародкового метамеру починає формуватися новий. Цей процес генетично детермінований та ритмічно повторюється багаторазово. При цьому метамер, утворений першим, розташовуватиметься в базальній частині втечі, а онтогенетично наймолодший - в апікальній. Накопичення числа зачатків метамерів у нирці Т.І. Серебрякова назвала її дозріванням. Для позначення максимальної кількості метамерів, що закладаються у нирці, вона запропонувала термін «ємність нирки».
Зростання та розвиток міжвузлів, як складових частин метамеру, значною мірою обумовлені діяльністю інтеркалярної меристеми. Розподіл клітин цієї меристеми та розтяг їх похідних призводить до подовження міжвузлів.
Метамери мають ряд характерних рис, які дозволяють їм забезпечувати структурну та функціональну цілісність рослинного організму.
1) Полярність метамерів. Кожен метамер має базальну та апікальну частину. Термін «апікальна частина» передбачає, що верхня частинаметамер або має у своєму складі апікальну меристему, або орієнтована в бік апексу.
Базальна та апікальна частини відрізняються за морфологічними, гістолого-анатомічними та фізіолого-біохімічними ознаками. Наприклад, у пшениці при переміщенні від базальної до апікальної частини міжвузля товщина стебла, діаметр медулярної лакуни і товщина периферичного кільця склеренхіми спочатку зростають, а потім поступово зменшуються; значно збільшується кількість клітин у складі тяжів асиміляційної паренхіми; зменшується радіальний діаметр провідних пучків, а також кількість судин у ксилемі.
2) Симетричність метамерів. Симетрія органів рослин забезпечується симетрією метамерів, що їх утворюють, яка виникає в результаті специфічної послідовності поділу клітин конуса наростання.
3) Гетерохронність утворення метамерів. Метамери різночасно, послідовно вичленюються конусом наростання. Тому перші метамери базальної частини втечі є онтогенетично старішими, а останні метамери апікальної частини – молодшими. Морфологічно вони відрізняються параметрами листа, довжиною та товщиною міжвузлів. Наприклад, у злаків листя верхнього ярусу ширше листя нижнього ярусу, а підколосові міжвузля довші і тонші за нижні.
4) Мінливість ознак метамерів. Онтогенетично молодші метамери мають меншу амплітуду мінливості ознак. Тому анатомічні ознаки підколосового міжвузля з більшою точністю можуть бути використані для ідентифікації сортів злакових культурта складання селекційних програм.
5) Оптимальність конструкції метамерів. Оптимальною біологічною конструкцією є така, на побудову та підтримку якої потрібна мінімальна кількість органічної речовини. Оскільки структура рослин нерозривно пов'язана з їхньою функцією, то критерієм оптимальності може бути пристосованість рослин до умов проростання та відношення їх насіннєвої продуктивності до маси вегетативних органів. Оптимальність конструкції цілісної рослини забезпечується оптимальністю конструкції складових його метамерів.
6) Цілісна рослина є полімерною системою, утвореною сукупністю метамерів. Полімерність проявляється усім рівнях організації рослинного організму. Так, у структурі ДНК є безліч повторень генів; каріотип багатьох рослин представлений диплоїдним чи поліплоїдним набором хромосом; у великій кількості у клітці зустрічаються пластиди, мітохондрії, рибосоми та інші органоїди. Доросла рослина є супідрядною сукупністю пагонів, коренів і репродуктивних органів, що мають метамерну будову. Метамерна система забезпечує високу продуктивність рослин і значно підвищує надійність їхнього онтогенезу.
27. 4.1. Первинна анатомічна будова кореня
Особливості первинної будови чітко виявляються на поздовжньому та поперечному зрізах кінчика кореня.
На поздовжньому зрізі кінчика кореня можна виділити чотири зони
Зона кореневого чохлика прикриває апікальні меристеми кореня. Вона складається із живих клітин. Їх поверхневий шар постійно злущується і вистилає хід, яким переміщається корінь. Клітини, що злущуються, крім того, виробляють слиз, який полегшує пересування кінчика кореня в грунті. Клітини центральної частини чохлика, або колумели, містять крохмальні зерна, що сприяє геотропному зростанню кореня. Клітини чохлика постійно оновлюються рахунок розподілу клітин спеціальної освітньої тканини – калітрогену, властивого однодольних.
I – зона кореневого чохлика; II – зона зростання; III – зона кореневих волосків; IV – зона проведення. 1 – епіблема, 2 – перицикл, 3 – ендодерма, 4 – первинна кора, 5 – екзодерма, 6 – центральний циліндр, 7 – кореневий волосок; 8 – формування бічного кореня.
Зона зростання і двох подзон. У підзоні поділу зростання кореня здійснюється завдяки активному мітотичному поділу клітин. Наприклад, у пшениці частка клітин, що діляться (мітотичний індекс) становить 100 – 200 проміле. Підзона розподілу є цінним матеріалом для цитогенетичних досліджень. Тут зручно вивчати число, макро- та мікроструктуру хромосом. У підзоні розтягування меристематична активність клітин знижується, але завдяки певному балансу фітогормонів, насамперед ауксинів та цитокінінів, зростання кореня відбувається за рахунок осьового розтягування молодих клітин.
Зона всмоктування з повним правом може бути названа зоною кореневих волосків, а також зоною диференціації, оскільки тут формуються епіблема, первинна кора та центральний циліндр.
Епіблема – це спеціальна покривна тканина, що постійно оновлюється, що складається з клітин двох типів. З трихобластів розвиваються тонкостінні кореневі волоски довжиною 1 – 3 мм, завдяки яким здійснюється всмоктування води та розчинених у ній речовин. Кореневі волоски недовговічні. Вони живуть 2 – 3 тижні, а потім злущуються. Атрихобласти кореневих волосків не утворюють та виконують покривну функцію.
Первинна кора регулює надходження води у провідні тканини центрального циліндра. При формуванні центрального циліндра першими утворюються тканини протофлоеми, якими в кінчик кореня надходять органічні речовини, необхідних його зростання. Вище за коренем лише на рівні перших кореневих волосків з'являються судинні елементи протоксилемы. Вихідні провідні тканини розвиваються в первинну флоему і первинну ксилему радіального пучка, що проводить.
Зона проведення займає найбільшу частину кореня. Вона виконує численні функції: транспорт води та розчинених у ній мінеральних і органічних речовин, синтез органічних сполук, запасання поживних речовин та інших. Зона проведення завершується кореневою шийкою, тобто. місцем переходу кореня до стебла. У дводольних покритонасінних на початку цієї зони відбувається перехід від первинного до вторинної анатомічної будови кореня.
На поперечному зрізі у зоні кореневих волосків виявляються структурні та топографічні особливості первинного анатомічного будови кореня (рис.2). На поверхні кореня знаходиться епіблема з кореневими волосками, за допомогою яких відбувається всмоктування води та розчинених у ній речовин. На відміну від епідермісу, епіблема не покрита кутикулою і не має продихів. Під нею розташовується первинна кора, що складається з екзодерми, мезодерми та ендодерми.
1 – зони кінчика кореня; 2 – 6 – поперечні зрізи різних рівнях; Вкс – вторинна ксилема; Вф - вторинна флоема; К – камбій; Мкс - метаксилема; Мф - метафлоема; Пд - протодерма; Пкс - протоксилема; Прд - Перідерма; Пф - протофлоема; Пц – перицикл; Рд - різодерма; Екз - екзодерма; Ен – ендодерма; Ч – чохлик.
Клітини екзодерми щільно складені та мають потовщені оболонки. Екзодерма забезпечує апопласне надходження води в глибші шари кори, а також надає кореню міцності з поверхні.
Середній шар кори, мезодерма, складається з тонкостінних паренхімних клітин і має пухке додавання завдяки наявності численних міжклітинників, якими вода переміщається до екзодерми.
Ендодерма, або внутрішній шар первинної кори, складається з ряду клітин. До її складу входять клітини з поясками Каспарі та пропускні клітини. Клітини з поясками Каспар мають потовщені бічні (радіальні) і тангентальні (торцеві) оболонки, звернені до центрального циліндра. Ці целюлозні потовщення просочуються лігніном і тому не можуть пропускати воду. На відміну від них пропускні клітини тонкостінні та розташовуються навпроти променів ксілеми. Саме через пропускні клітини вода надходить у радіальний провідний пучок центрального циліндра.
Центральний циліндр кореня, або стела, складається з кількох шарів клітин. Відразу під ендодермою знаходиться один або кілька рядів клітин перициклу. З них при переході до вторинної будови кореня у дводольних покритонасінних та голонасінних утворюються міжпучковий камбій та феллоген (пробковий камбій). Крім того клітини перициклу беруть участь в утворенні бічних коренів. За перициклом розташовуються клітини радіального пучка, що проводить.
Пучок, що проводить, формується з прокамбію. Спочатку утворюються клітини протофлоеми, та був лише на рівні перших кореневих волосків – клітини протоксилемы. Клітини які проводять тканин з'являються екзархно, тобто. з поверхні пучка, і надалі розвиваються в доцентровому напрямку. І тут найперші, більші судини перебувають у центрі ксилемы, а молодші, меншого діаметра – на периферії ксилемного променя. Між променями ксілеми розташовані клітини флоеми.
Число променів ксілеми залежить від систематичного становища рослин. Наприклад, у деяких папоротей може бути лише один промінь ксілеми та одна ділянка флоеми. Тоді пучок називається монархним. Для багатьох дводольних покритонасінних характерні діархні пучки з двома променями ксилеми. Крім того, у них зустрічаються рослини з три-, тетра- та пентархними пучками. У однодольних покритонасінних типові поліархні провідні пучки з багатопроменевою ксилемою.
Будова радіального провідного пучка впливає спосіб закладання бічних коренів. У діархному корені вони формуються між флоемою та ксилемою, у триархному та тетрархному – навпроти ксилеми, а в поліархному – навпроти флоеми.
Клітинна будова епіблеми та первинної кори забезпечує виникнення кореневого тиску води. Вода всмоктується кореневими волосками, їх виходить у клітинні оболонки екзодерми, потім надходить у межклетники мезодерми, та якщо їх через тонкостінні пропускні клітини – в судини радіального провідного пучка. Оскільки кореневі волоски мають більшу сумарну поверхню, ніж пропускні клітини, то швидкість пересування води, отже, та її тиск зростають у міру наближення до судин. Різниця тисків, що виникають, становить кореневий тиск, який є одним з механізмів надходження води в стебло та інші органи рослин. Другим важливим механізмом пересування води є транспірація.
28. 4.2. Вторинна анатомічна будова кореня
У дводольних покритонасінних та голонасінних первинна анатомічна будова кореня в зоні проведення доповнюється структурами вторинного походження, які формуються завдяки виникненню та меристематичній активності вторинних бічних освітніх тканин – камбію та феллогену (коркового камбію) (рис. 3). При переході до вторинної будови докорінно відбуваються такі істотні зміни.
А. Поява в центральному циліндрі кореня камбію і породжуваних ним вторинної ксилеми і вторинної флоеми, які за пропускною здатністю набагато перевершують елементи вихідного радіального пучка, що проводить.
Цей тип відноситься до складних тканин, складається з по-різному диференційованих клітин. Крім власне провідних елементів, у тканині присутні механічні, видільні та запасні елементи. Провідні тканини поєднують всі органи рослини єдину систему. Виділяють два типи провідних тканин: ксилему та флоему (грец.xylon – дерево; phloios – кора, лико). Вони мають як структурні, і функціональні відмінності.
Провідні елементи ксилеми утворені мертвими клітинами. По них здійснюється далекий транспорт води та розчинених у ній речовин від кореня до листя. Провідні елементи флоеми зберігають живий протопласт. По них здійснюється далекий транспорт від фотосинтезуючого листя до кореня.
Зазвичай ксилема і флоема розташовуються в тілі рослини в певному порядку, утворюючи шари або пучки, що проводять. Залежно від будови розрізняють кілька типів пучків, що проводять, які характерні для певних груп рослин. У колатеральному відкритому пучку між ксилемою та флоемою знаходиться камбій, що забезпечує вторинне зростання. У біколатеральному відкритому пучку флоема розташовується щодо ксилеми з двох сторін. Закриті пучки не містять камбію, а звідси до вторинного потовщення не здатні. Можна зустріти ще два типи концентричних пучків, де або флоема оточує ксилему, або ксилема – флоему.
Ксилема (деревина). Розвиток ксілеми у вищих рослин пов'язане із забезпеченням водного обміну. Оскільки через епідерму постійно виводиться вода, така ж кількість вологи має поглинатися рослиною і додаватися до органів, які здійснюють транспірацію. Слід враховувати, що наявність живого протопласту в клітинах, що проводять воду, сильно сповільнювало б транспорт, мертві клітини тут виявляються більш функціональними. Однак мертва клітина не має тургесцентності, у зв'язку з цим механічними властивостямиповинна мати оболонка. Примітка: тургесценція - стан рослинних клітин, тканин та органів, при кото?и? вони стають пружними внаслідок тиску вмісту клітин з їхньої еластичні оболонки. Дійсно, провідні елементи ксилеми складаються їх витягнутих вздовж осі органу мертвих клітин з товстими оболонками, що одеревіли.
Спочатку ксилема утворюється з первинної меристеми – прокамбію, розташованого на верхівках осьових органів. Спочатку диференціюється протоксилема, потім метаксилема. Відомо три типи формування ксилеми. При екзархному типі елементи протоксилем спочатку з'являються на периферії пучка прокамбію, потім в центрі виникають елементи метаксилеми. Якщо процес йде у протилежному напрямку (тобто від центру до периферії), це ендархний тип. При мезархном типі ксилема закладається в центрі прокамбіального пучка, після чого відкладається як до центру, так і до периферії.
Для кореня характерний екзархний тип закладки ксілеми, для стебел – ендархний. У низькоорганізованих рослин способи формування ксилемы дуже різноманітні можуть служити систематичними характеристиками.
Деякі? рослин (наприклад, однодольних) всі клітини прокамбію диференціюються у провідні тканини, які не здатні до вторинного потовщення. В інших форм (наприклад, деревних) між ксилемою і флоемою залишаються латеральні меристеми (камбій). Ці клітини здатні ділитися, оновлюючи ксилему та флоему. Такий процес називається вторинним зростанням. У багатьох, що виростають у порівняно стабільних кліматичних умовах, рослин, зростання йде постійно. У форм, пристосованих до сезонних змін клімату, періодично.
Основні етапи диференціації клітин прокамбію. Її клітини з тонкими оболонками, що не перешкоджають їх розтягуванню при зростанні органу. Потім протопласт починає відкладати вторинну оболонку. Але це процес має виражені особливості. Вторинна оболонка відкладається не суцільним шаром, що дозволило б клітині розтягуватися, а вигляді кілець чи з спіралі. Подовження клітини у своїй не утруднено. У молодих клітин кільця чи витки спіралі розташовані близько друг до друга. У зрілих клітин розходяться внаслідок розтягування клітини. Кільчасті та спіральні потовщення оболонки росту не перешкоджають, проте механічно вони поступаються оболонкам, де вторинне потовщення утворює суцільний шар. У зв'язку з цим після припинення зростання в ксилемі формуються елементи з суцільною здерев'янілою оболонкою (метаксилемою). Слід зазначити, що вторинне потовщення тут не кільчасте або спіральне, а точкове, сходове, сітчасте. Її клітини розтягуються, не здатні протягом кількох годин відмирають. Цей процес у розташованих поблизу клітин відбувається скоординовано. У цитоплазмі утворюється велика кількість лізосом. Потім розпадаються, а ферменти, що знаходяться в них, руйнують протопласт. При руйнуванні поперечних стінок розташовані ланцюжком одна над одною клітини утворюють порожнисту судину. Більшість покритонасінних рослин і деякі? папоротьподібні мають судини.
Клітку, що проводить, не утворює наскрізних перфорацій у своїй стінці, називають трахеїдою. Пересування води трахеїдами йде з меншою швидкістю, ніж судинами. Справа в тому, що у трахеїдів ніде не переривається первинна оболонка. Між собою трахеїди повідомляться за допомогою пір. Слід уточнити, що у рослин пора є лише поглиблення у вторинній оболонці до первинної оболонки і ніяких наскрізних перфорацій між трахеїдами немає.
Найчастіше зустрічаються облямовані пори. Вони канал, звернений у порожнину клітини, утворює розширення - камеру пори. Пори більшості хвойних рослинна первинній оболонці мають потовщення - торус, який є своєрідним клапаном і здатний регулювати інтенсивність транспорту води. Зміщуючись, торус перекриває струм води через пору, але після цього повернутися в колишнє положення він не може, роблячи одноразове дію.
Пори бувають більш менш округлими, витягнутими перпендикулярно витягнутої осі (група цих пір нагадує сходи, у зв'язку з цим таку пористість називають сходовою). Через пори транспорт здійснюється як у поздовжньому, так і у поперечному напрямку. Пори присутні не тільки у трахеїд, а й окремих клітин судин, які утворюють судину.
З погляду еволюційної теорії трахеїди є першою та основною структурою, що здійснює проведення води в тілі вищих рослин. Вважають, що судини виникли з трахеїд внаслідок лізису поперечних стінок між ними. Більшість папоротеподібних та голонасінних судин не мають. Пересування води у них відбувається за допомогою трахеїдів.
У процесі еволюційного розвитку судини виникали у різних групрослин неодноразово, але найбільш важливе функціональне значення вони набули у покритонасінних, у кото?и? вони є поряд із трахеїдами. Вважають, що володіння більш досконалим механізмом транспорту допомогло їм не лише вижити, а й досягти значного розмаїття форм.
Ксилема є складною тканиною, крім водопровідних елементів у ній містяться інші. Механічні функціївиконують волокна лібриформа (лат. liber – луб, forma – форма). Присутність додаткових механічних структур важливо, оскільки, незважаючи на потовщення, стінки водопровідних елементів все ж таки занадто тонкі. Вони не здатні самостійно утримувати велику масу багаторічної рослини. Волокна розвивалися із трахеїд. Їх характерні менші розміри, одеревенілі (лігніфіковані) оболонки і вузькі порожнини. На стіні можна виявити, позбавлені облямівки пори. Ці волокна проводити воду не можуть, основна їхня функція опорна.
У ксілемі є живі клітини. Їхня маса може досягати 25% від загального обсягу деревини. Оскільки ці клітини мають округлу форму, їх називають паренхімою деревини. У тілі рослини паренхіма розташовується двома способами. У першому випадку клітини розташовуються у вигляді вертикальних тяжів – це тяжова паренхіма. В іншому випадку паренхіма утворює горизонтальні промені. Вони називаються сердечними променями, оскільки з'єднують серцевину і кору. Серцевина виконує низку функцій, зокрема і запасання речовин.
Флоема (луб). Це складна тканина, оскільки утворена різнотипними клітинами. Основні клітини, що проводять, називаються ситовидними елементами. Провідні елементи ксилеми утворені мертвими клітинами, а у флоеми вони протягом періоду функціонування зберігають живий, хоч і сильно змінений протопласт. По флоемі відбувається відтік пластичних речовин від фотосинтезуючих органів. Здатність проводити органічні речовини мають усі живі клітини рослин. А звідси, якщо ксилему можна знайти лише у вищих рослин, то транспорт органічних речовин між клітинами здійснюється і в нижчих рослин.
Ксилема та флоема розвиваються з апікальних меристем. На першому етапі у прокамбіальному тяжі формується протофлоема. У міру зростання оточуючих тканин вона розтягується, і коли зростання завершується, замість протофлоеми формується метафлоема.
У різних груп вищих рослин можна зустріти два типи ситоподібних елементів. У папоротеподібних і голонасінних він представлений ситоподібними клітинами. Ситоподібні поля у клітинах розсіяні з бокових стінок. У протопласті зберігається деструктоване ядро.
У покритонасінних ситоподібні елементи називаються ситоподібними трубками. Вони повідомляються між собою через ситоподібні платівки. У зрілих клітинах ядра відсутні. Однак поруч із ситовидною трубкою розташовується клітина-супутниця, що утворюється разом із ситовидною трубкою в результаті мітотичного поділу загальної материнської клітини (рис.38). Клітина-супутниця має більш щільну цитоплазму з великою кількістю активних мітохондрій, а також повноцінно функціонуюче ядро, величезну кількість плазмодесм (вдесятеро більше, ніж в інших клітин). Клітини-супутниці впливають на функціональну активність без'ядерних ситовидних клітин трубок.
Структура зрілих ситоподібних клітин має деякі особливості. Відсутня вакуоля, у зв'язку з цим цитоплазма сильно розріджується. Може бути відсутнім (у покритонасінних рослин) або перебувати в зморщеному функціонально малоактивному стані ядро. Рибосоми та комплекс Гольджі також відсутні, але добре розвинений ендоплазматичний ретикулум, який не тільки пронизує цитоплазму, а й переходить до сусідніх клітин через пори ситоподібних полів. Добре розвинені мітохондрії та пластиди зустрічаються удосталь.
Між клітинами транспорт речовин йде через отвори, що розташовані на клітинних оболонках. Такі отвори називаються порами, але на відміну від пір трахеїд є наскрізними. Припускають, що вони є сильно розширеними плазмодесми, на стінках, які? відкладається полісахарид калозу. Пори розташовуються групами, утворюючи ситоподібні поля. У примітивних форм ситоподібні поля безладно розсіяні по всій поверхні оболонки, у більш досконалих покритонасінних рослин розташовуються на сусідніх клітин, що примикають один до одного, утворюючи ситовидну пластинку. Якщо на ній знаходиться одне ситоподібне поле, її називають простою, якщо дещо складною.
Швидкість пересування розчинів по ситоподібних елементах становить до 150см за годину. Це у тисячу разів перевищує швидкість вільної дифузії. Ймовірно, має місце активний транспорт, а численні мітохондрії ситоподібних елементів та клітин-супутниць постачають для цього необхідну АТФ.
Термін діяльності ситоподібних елементів флоеми залежить від наявності латеральних меристем. Якщо вони є, то ситоподібні елементи працюють протягом усього життя рослини.
Крім ситоподібних елементів і клітин-супутниць, у флоемі присутні луб'яні волокна, склереїди та паренхіма.
У процесі еволюції з виходом вищих рослин на сушу вони виникли тканини, які досягли своєї найбільшої спеціалізації у квіткових рослин. У цій статті ми розглянемо докладніше, що є тканини рослин, які види їх існують, які функції вони виконують, а також особливості будови тканин рослин.
Тканину називають групи клітин, подібних за своєю будовою та виконують однакові функції..
Основні тканини рослин представлені на малюнку нижче:
Види, функції та будова тканин рослин.
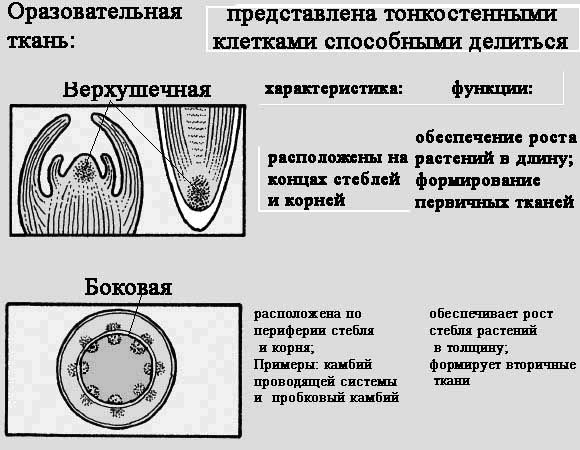

Покривна тканина рослин.


Покривна тканина рослин - кірка
Тканина рослин, що проводить.
| Назва тканини | Будова | Місцезнаходження | Функції |
| 1. Судини деревини – ксилема | Порожні трубки з стінами, що одеревіють, і відмерлим вмістом | Деревина (ксилема), що проходить уздовж кореня, стебла, жилок листя | Проведення води та мінеральних речовин із ґрунту в корінь, стебло, листя, квітки |
|
2.Ситовидні трубки лубу - флоема Супроводжуючі клітини або клітини-супутниці |
Вертикальний ряд живих клітин із ситоподібними поперечними перегородками Сестринські клітини ситоподібних елементів, що зберегли свою структуру |
Луб (флоема), розташований уздовж кореня, стебла, жилок листя Завжди розташовуються вздовж ситоподібних елементів (супроводжують їх) |
Проведення органічних речовин із листя в стебло, корінь, квітки Приймають активну участь у проведенні органічних речовин з ситоподібних трубок флоеми |
| 3. Провідні судинно-волокнисті пучки | Комплекс з деревини та лубу у вигляді окремих тяжів у трав та суцільного масиву у дерев | Центральний циліндр кореня та стебла; жилки листя та квіток | Проведення з деревини води та мінеральних речовин; по лубу – органічних речовин; зміцнення органів, зв'язок їх у єдине ціле |



Механічна тканина рослин.

